第5章 感覚と知覚

図5.1 | もしあなたがこの街角の風景の中に立っていたとしたら、あなたは数多くの感覚的な情報を吸収し、処理していることでしょう。(credit: modification of work by Cory Zanker)
この章の概要
5.1 感覚と知覚
5.2 波と波長
5.3 視覚
5.4 聴覚
5.5 その他の感覚
5.6 知覚についてのゲシュタルト原理
はじめに
都会の街角に立っているところを想像してみてください。あなたは、あちらこちらにいる目的地へ向かう車や人の動き、ストリートミュージシャンのメロディーや遠くで鳴らされているクラクションの音、排気ガスやそばの路上販売で売られている食べ物の匂い、そして足元の硬い舗装の感触など、さまざまなものを受け取ります。
私たちは、周囲の環境についての重要な情報を得るために、感覚系に依拠しています。私たちはこの情報を使って環境をうまく切り抜けたり環境と相互作用したりすることで、食糧を見つけ、住処を探し、社会的な関係を維持し、そして潜在的に危険な状況を回避します。
この章では、感覚情報が神経系によってどのように受け取られ、処理されるのか、そしてそれが私たちの意識的な世界の経験にどのように影響するのかを概観します。私たちはまず、感覚と知覚の違いを学ぶことから始めます。続いて、私たちは光と音の刺激の物理的な性質と、主要な感覚系の基本的な構造と機能の概要を検討します。この章は、ゲシュタルトと呼ばれる歴史的に重要な知覚理論の議論で締めくくられます。
5.1 感覚と知覚
学習目標
この節が終わるまでに、あなたは次のことができるようになります:
- 感覚と知覚を区別する
- 絶対閾値と弁別閾値の概念を記述する
- 知覚における注意、動機付け、感覚適応の役割を議論する
感覚
何かを感じるということはどういうことでしょうか?感覚受容体は、特定の種類の刺激に反応する特殊なニューロンです。感覚情報が感覚受容体によって検出されると、感覚が生じます。たとえば、目に入った光は、目の奥にある細胞に化学変化を起こします。これらの細胞は、(生物心理学を学習する際に学んだように)活動電位の形でメッセージを中枢神経系に伝えます。感覚刺激のエネルギーが活動電位に変換されることは、伝達として知られています。
あなたはおそらく、私たちが視覚、音(聴覚)、匂い(嗅覚)、味(味覚)、感触(体性感覚)という5つの感覚を有していることは小学生の頃から知っていることでしょう。しかしながら、この五感という概念は単純化されすぎていることがわかっています。私たちはまた、平衡(前庭感覚)、体の位置や動き(固有受容感覚と運動感覚)、痛み(侵害受容感覚)、および温度(熱受容感覚)についての情報を提供する感覚系も有しています。
関連する刺激に対する所与の感覚系の感度は、絶対閾値として表すことができます。絶対閾値とは、その刺激が50%の確率で検出されるために存在しなければならない、刺激エネルギーの最小量のことを指します。これを考える別の方法とは、2回に1回検出されるためには光をどれだけ暗くすることができるか、あるいは音をどれだけ小さくすることができるか、を尋ねるというものです。私たちの感覚受容体の感度はまったく驚くべきものです。晴れた夜には、目の奥にある最も感度の高い感覚細胞は、30マイル[約48キロメートル]先のろうそくの炎を感知できると推定されています(Okawa & Sampath, 2007)。静かな環境下では、有毛細胞(内耳の受容体細胞)は20フィート[約6メートル]先の時計の音を検知することができます(Galanter, 1962)。
また、私たちは意識的に認識できる閾値を下回る形で与えられるメッセージを受け取ることも可能です。これらはサブリミナル・メッセージと呼ばれます。ある刺激が生理学的な閾値に達するのは、その刺激が感覚受容体を励起させ、神経インパルスを脳に送るのに十分な強さであるときです:これが絶対閾値です。この閾値よりも下のメッセージはサブリミナルと呼ばれます:私たちはそれを受け取りますが、私たちはそれについて意識的に気づいていません。広告やロックミュージック、および自己啓発のオーディオ・プログラムにおけるサブリミナル・メッセージの使用について、長年にわたりさまざまな憶測が飛び交ってきました。研究の証拠によれば、実験室の環境においては、人々は認識の外側にある情報を処理し、反応することができるということが示されています。しかし、これは私たちがそれらのメッセージにゾンビのように従うということを意味しているのではありません。実際、隠されたメッセージは、実験室の外では行動にほとんど影響を与えません(Kunst-Wilson & Zajonc, 1980; Rensink, 2004; Nelson, 2008; Radel, Sarrazin, Legrain, & Gobancé, 2009; Loersch, Durso, & Petty, 2013)。
絶対閾値は一般的に、感度にとって最適な状況において、信じられないほど制御された条件下で測定されます。時には、私たちは2つの刺激の間の違いを検出するために、どれだけの差異が必要かということに関心があります。これは丁度可知差異(jnd)または弁別閾値として知られています。絶対閾値とは異なり、弁別閾値は刺激の強度に基づいて変化します。一例として、自分が真っ暗な映画館の中にいるところを想像してみてください。もし、観客の1人がテキストメッセージを受信して携帯電話の画面が光ったとしたら、多くの人が映画館の中の明かりの変化に気づくでしょう。しかしながら、もし同じことがバスケットボールの試合中の煌々と照らされたアリーナで起こったとしたら、気づく人はほとんどいないでしょう。携帯電話の明るさは変わっていませんが、明かりの変化として検知される能力が、2つの文脈では劇的に異なっています。エルンスト・ヴェーバーは1830年代にこの弁別閾値の変化の理論を提唱し、それはヴェーバーの法則として知られるようになりました:ヴェーバーの法則とは、弁別閾値は元の刺激の一定の割合のものである(上の例で示されるように)、ということです。
知覚
私たちの感覚受容体は常に環境からの情報を収集していますが、私たちがどのようにして世界と相互作用するかに影響を与えるのは、究極的には私たちがその情報をどのようにして解釈するかということです。知覚とは、感覚情報が整理され、解釈され、意識的に経験される方法を指します。知覚には、ボトムアップ処理とトップダウン処理があります。図5.2に示されるように、ボトムアップ処理とは、環境中の刺激からの感覚情報が処理を駆動することを意味し、トップダウン処理とは、知識と期待が処理を駆動することを意味します(Egeth & Yantis, 1997; Fine & Minnery, 2009; Yantis & Egeth, 1999)。
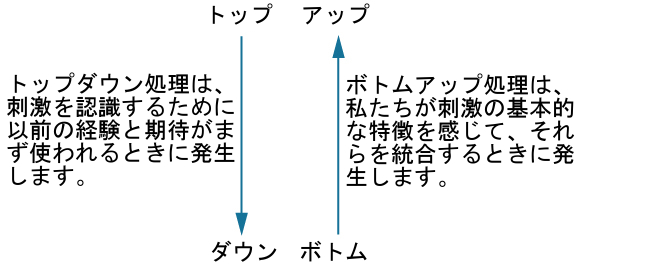
図5.2 | トップダウンとボトムアップは、私たちが知覚を処理する方法です。
あなたと何人かの友人が、混雑したレストランの中に座って昼食をとりながら話をしていると想像してみてください。とても騒がしいので、あなたは友人が何を話しているのかを聞き取ろうと彼女の顔に集中していると、ガラスの割れる音や金属製の皿が床にぶつかる音が鳴り響いてきます。接客係が料理の入った大きなトレイを落としたのです。あなたが食事や会話に気を配っていたとしても、その衝撃音は注意のフィルターを通り抜けて、あなたの注意を引くことになるでしょう。あなたは気づかずにはいられません。このような注意の引きつけ方は、環境からの音によって引き起こされるものであり、ボトムアップとなります。
一方、トップダウン処理は、一般的には目標に向けられた、ゆっくりとして、意図的で、努力のいる、自分のコントロール下にあるものです(Fine & Minnery, 2009; Miller & Cohen, 2001; Miller & D’Esposito, 2005)。たとえば、あなたは鍵を紛失した場合、どのようにして探しますか?もしあなたが黄色いキーホルダーをつけていたとしたら、おそらくあなたはカウンターやコーヒーテーブル、その他の似たような場所といった特定の場所で、一定の大きさの黄色いものを探すでしょう。あなたはシーリングファンの上で黄色いものを探すことはないでしょう。なぜなら、普通、鍵はシーリングファンの上には転がっていないということをあなたは知っているからです。このように、ある場所において一定の大きさの黄色いものを探し、他の場所では探さないという行為は、トップダウンです。それらは、あなたのコントロール下にあり、あなたの経験に基づいています。
この概念について考える1つの方法は、感覚が物理的なプロセスであるのに対し、知覚は心理的なプロセスであると考えることです。たとえば、キッチンに入ってシナモンロールを焼いている香りを嗅いだとき、感覚としては香りの受容体がシナモンの香りを感知していますが、知覚としては「うーん、これは休日に家族が集まったときにおばあちゃんが焼いていたパンの香りみたいだな」というものかもしれません。
私たちの知覚は感覚から成り立っていますが、すべての感覚が知覚につながるわけではありません。実際、長期間にわたって比較的一定の刺激を受け続けている場合、私たちはしばしばそれを知覚しなくなります。これは、感覚適応として知られています。あなたが一度も訪れたことのない街に行ったところを想像してみてください。あなたはホテルにチェックインしますが、部屋に着くと、窓の外には道路工事の看板があり、明るく点滅しています。残念ながら他の部屋は空いていないので、あなたは点滅する光に悩まされることになります。あなたはテレビを見てくつろぐことにします。あなたが最初に部屋に入ったときには、点滅する光が非常に気になりました。それはまるで誰かが部屋の中で明るい黄色のスポットライトを点けたり消したりし続けているかのようでしたが、しばらくテレビを見ているうちに、あなたは光の点滅に気づかなくなりました。相変わらず光が数秒ごとに点滅して部屋中が黄色い光で照らされており、あなたの目の中の光受容体はまだその光を感じていますが、あなたはもはや光の状態の急激な変化を知覚していません。あなたが光の点滅を知覚しなくなったということは、感覚適応を示しています。それは、感覚と知覚とは密接に関連しているものの、異なるものであることを示しています。
感覚と知覚に影響を与えるもう1つの要素があります:それは、注意です。注意は、何が感じられるかと、何が知覚されるかを決定する上で重要な役割を果たします。あなたが、音楽やおしゃべり、笑い声の飛び交うパーティーに参加しているところを想像してください。あなたは1人の友人との興味深い会話に夢中になり、周囲の雑音をすべて排除しています。もし、誰かが会話に入ってきて、今かかっていた曲は何かと尋ねてきたら、あなたはおそらくその質問に答えられないでしょう。
学習へのリンク
シモンズとチャブリス(Simons and Chabris, 1999)によるこの選択的注意テスト(http://openstax.org/l/blindness)を試してみることによって、非注意性盲目がどのように作用するかを自分の目で確かめてみましょう。
私たちが環境を知覚する上で、注意がいかに重要であるかを示す最も興味深い研究の1つに、ダニエル・シモンズとクリストファー・チャブリスが行った有名な研究があります(Simons and Chabris, 1999)。この研究では、白い服と黒い服を着た人がバスケットボールをパスしている映像を参加者に見せました。参加者には、白い服を着たチームがボールをパスした回数を数えてもらいました。ビデオの途中で、黒いゴリラの着ぐるみを着た人が、2つのチームの間を歩いていきます。あなたは人々がゴリラに気づくと思いますよね?しかし、9秒間はっきりとゴリラが見えていたという事実にもかかわらず、ビデオを見た人のうち半数近くがゴリラに全く気づかなかったのです。参加者は白い服を着たチームがボールをパスする回数に非常に集中していたため、彼らは他の視覚情報を完全に無視していたのです。非注意性盲目とは、人が何かに積極的に関心を向けており、その他のことに注意を払っていないために、完全に見えている何かに気づくことができないことです(Mack & Rock, 1998; Simons & Chabris, 1999)。
同様の実験で、研究者は、参加者に対しコンピュータ画面上を移動する画像を観察するように指示することにより、非注意性盲目をテストしました。参加者は、白色または黒色の物体に注目し、他の色は無視するように指示されました。赤色の十字が画面を横切ったとき、約3分の1の被験者はそれに気づきませんでした(図5.3)(Most, Simons, Scholl, & Chabris, 2000)。

図5.3 | ある研究の参加者の約3分の1は、注意を黒色や白色の図形に集中させていたため、赤色の十字架がスクリーン上を通過したことに気づきませんでした。(credit: Cory Zanker)
動機付けも知覚に影響を与えることがあります。あなたは、本当に大切な電話を待っていて、シャワーを浴びているときに電話が鳴ったと思ったら、実は鳴っていなかったという経験はありませんか?もしあるなら、あなたは、意味のある刺激を検出したいという動機付けが、本当の感覚刺激と背景の雑音とを識別する能力を変化させるのを経験したことになります。このように、刺激が紛らわしい背景に埋め込まれているときでも、それを識別する能力のことを信号検出理論といいます。これは、母親が赤ん坊の静かなつぶやきで目を覚ますのに、眠っている間に発生する他の音では目を覚まさない理由も説明してくれるかもしれません。信号検出理論は、航空管制官の正確性向上などに実践的な応用があります。航空管制官は、レーダー画面に現れる多くの信号(ブリップ)の中から飛行機を検出し、それらの飛行機が空を移動するのを追跡することができる必要があります。実際、信号検出理論を開発した研究者の元々の研究は、飛行機のブリップに対する航空管制官の感度を向上させることに焦点を当てたものでした(Swets, 1964)。
私たちの知覚は、私たちの信念、価値観、先入観、期待、そして人生経験によっても影響を受けることがあります。あなたがこの章の後半で見ていくことになるように、発達の重要な時期に両眼視の経験を奪われた人は、奥行きを知覚するのが難しくなります(Fawcett, Wang, & Birch, 2005)。ある文化的背景の中で人々が共有する経験は、知覚に顕著な影響を与えます。たとえば、マーシャル・セガール、ドナルド・キャンベル、メルヴィル・ハースコヴィッツ(Segall, Campbell, and Herskovits, 1963)は、多国間の研究結果を発表しました。その中で彼らは、西洋文化圏の人は非西洋文化圏の人に比べて、特定のタイプの視覚的な錯覚を経験しやすく、その逆もまた然りであることを示しました。西洋人がより経験しやすい錯視の1つに、ミューラー・リヤー錯視(図5.4)があります:これらの線は長さが違うように見えますが、実際には同じ長さです。
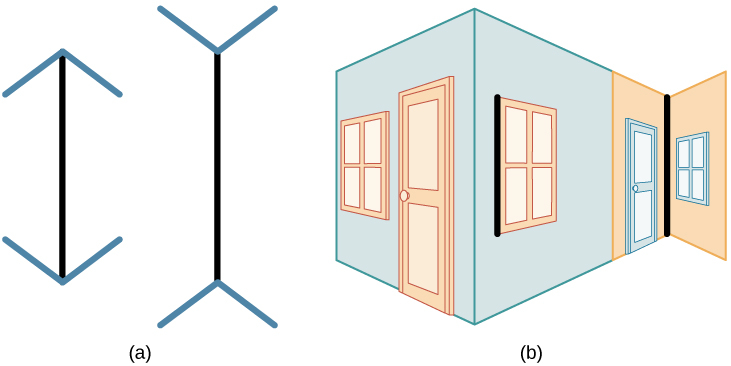
図5.4 | ミューラー・リヤー錯視では、2本の線の長さは同じ長さであるものの、それらは違って見えます。(a)線の端に矢印があると、同じ長さの線であっても、右側の線が長く見えることがあります。(b)三次元画像に当てはめると、黒い線は2本とも同じ長さですが、やはり右側の線が長く見えることがあります。
このような知覚の違いは、ある文化的な文脈を持つ人々が日常的に経験している環境特性の種類の違いと一致していました。たとえば、西洋文化圏の人々は、直線で構成された建物という知覚的な文脈を持っており、セガールの研究ではこれを「大工世界」と呼びました(Segall et al., 1966)。一方、丸い小屋を円形に配置して村を形成している南アフリカのズールー族のような、非大工的な見方を持つ特定の非西洋文化圏の人々は、このような錯覚に陥りにくいです(Segall et al., 1999)。文化的な要因によって影響を受けるのは、視覚だけではありません。実際、匂いを識別し、その心地よさや強さを評価する能力は、文化によって異なることが研究によって実証されています(Ayabe-Kanamura, Saito, Distel, Martínez-Gómez, & Hudson, 1998)。
また、スリルを求める子供は、強烈な酸味への好みをより示す傾向があります(Liem, Westerbeek, Wolterink, Kok, & de Graaf, 2004)。これは、人格の基本的な側面が知覚に影響を与えている可能性を示唆しています。さらに、低脂肪食品に対して肯定的な態度をとっている人は、これらの製品についてあまり肯定的な態度をとっていない人に比べて、低脂肪と表示された食品をより美味しく感じると評価する傾向があります(Aaron, Mela, & Evans, 1994)。
5.2 波と波長
学習目標
この節が終わるまでに、あなたは次のことができるようになります:
- 波の形態の重要な物理的特性を記述する
- 光波の物理的特性が知覚経験とどのように関連しているかを示す
- 音波の物理的特性が知覚経験とどのように関連しているかを示す
視覚と聴覚の刺激は、どちらも波の形態で起こります。この2つの刺激は構成の上では非常に異なっていますが、波の形態は、私たちの視覚と聴覚の知覚にとって特に重要な、類似の特徴を共有しています。この節では、私たちは、波の物理的特性と、それに関連する知覚経験について記述します。
振幅と波長
波の物理的特性には、振幅と波長があります(図5.5)。波の振幅とは、中心線から山の頂点または谷の底の点までの距離のことです。波長とは、ある頂点から次の頂点までの波の長さのことを指します。

図5.5 | 波の振幅(すなわち高さ)は、山までの長さ、あるいは谷までの長さから測定されます。波長は頂点から頂点までの長さから測定されます。
波長は、ある波形の周波数に直接関係します。周波数とは、所与の期間内にある地点を通過する波の数のことで、しばしばヘルツ(Hz)、つまり1秒あたりのサイクル数で表されます。波長が長ければ周波数は低く、波長が短ければ周波数は高くなります(図5.6)。

図5.6 | この図は、異なる波長/周波数の波を表しています。図の一番上にある赤色の波は、波長が長く、周波数が低いです。上から下に向かうにつれて、波長は短くなり、周波数は高くなっています。
光波
可視スペクトルは、より大きな電磁スペクトルのうち、私たちが目で見ることのできる部分です。図5.7が示すように、電磁スペクトルは、ガンマ線、X線、紫外光、可視光、赤外光、マイクロ波、電波など、私たちの環境で生じるすべての電磁放射を含んでいます。人間の可視スペクトルは、380~740nm(ナノメートル(nm)は10億分の1メートルであるため、非常に小さな距離です)の範囲の波長に関連しています。他の生物種は、電磁スペクトルの他の部分を検出することができます。たとえば、ミツバチは紫外域の光を見ることができ(Wakakuwa, Stavenga, & Arikawa, 2007)、一部のヘビは、従来の視覚的な光の手がかりに加えて、赤外放射を検出することができます(Chen, Deng, Brauth, Ding, & Tang, 2012; Hartline, Kass, & Loop, 1978)。

図5.7 | 人間が見ることのできる光は、電磁スペクトルのごく小さな一部を構成するにすぎません。
人間の場合、光の波長は色の知覚と関連しています(図5.8)。可視スペクトルの中では、私たちの赤色の経験はより長い波長に関連付けられており、緑色は中間の波長、青色や紫色は波長が短いです。これを簡単に覚える方法は、ROYGBIVという暗記法です:赤色(red)、橙色(orange)、黄色(yellow)、緑色(green)、青色(blue)、藍色(indigo)、紫色(violet)。また、光波の振幅は、明るさや色の強さについての私たちの経験と関連しており、振幅が大きいほど明るく見えます。
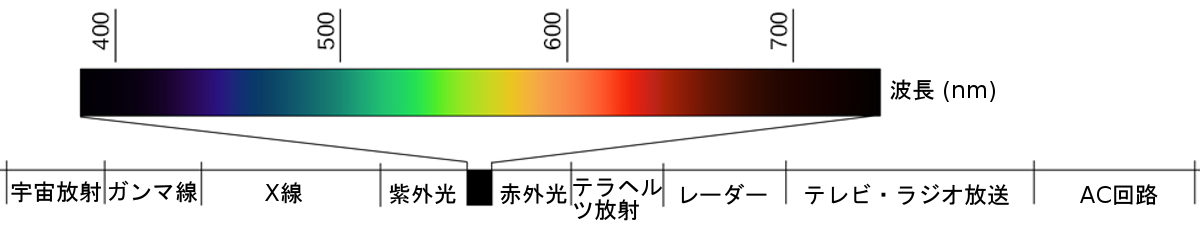
図5.8 | 光の異なる波長は、異なる色についての私たちの知覚に関連しています。(credit: modification of work by Johannes Ahlmann)
音波
光波と同様に、音波の物理的特性は、私たちの音の知覚のさまざまな側面と関連しています。音波の周波数は、その音の高さについての私たちの知覚と関連しています。高い周波数の音波は高い音として知覚され、低い周波数の音波は低い音として知覚されます。音の周波数の可聴範囲は20~20000Hzであり、この範囲の中央に位置する周波数に対して最も感度が高いです。
可視スペクトルの場合と同様に、他の生物種は異なる可聴範囲を示します。たとえば、ニワトリは125~2000Hzと非常に限られた可聴域を有しています。ネズミの可聴域は1000~91000Hz、シロイルカの可聴域は1000~123000Hzです。私たちが飼っている犬や猫の可聴域は、それぞれ70~45000Hz、45~64000Hz程度です(Strain, 2003)。
ある音の大きさは、音波の振幅と密接に関係しています。より大きな振幅は、より大きな音に関連付けられます。音の大きさは、音の強さを表す対数の単位であるデシベル(dB)で測定されます。典型的な会話であれば60dB、ロックコンサートであれば120dBとなるでしょう(図5.9)。5フィート先のささやき声や木の葉のそよぐ音は、私たちの可聴範囲の下限に位置します。窓のエアコンの音、普通の会話、さらには激しい車の行きかいや掃除機の音でも、許容範囲内です。しかしながら、約80dBから130dBの範囲では、聴覚に損傷を与える可能性があります:これは、フードプロセッサー、芝刈り機、大型トラック(25フィート[約8メートル]先)、地下鉄(20フィート[約6メートル]先)、ロック音楽のライブ、削岩機などの音です。聴力喪失の約3分の1は騒音への曝露によるもので、音が大きければ大きいほど、聴覚障害を引き起こすのに必要な曝露時間は短くなります(Le, Straatman, Lea, & Westerberg, 2017)。最大音量(約100~105デシベル)でイヤホンを使って音楽を聴くと、15分間の曝露で騒音性聴力喪失を引き起こすことがあります。最大音量で音楽を聴いても損傷を受けないように見えますが、加齢による聴力喪失のリスクが高まります(Kujawa & Liberman, 2006)。痛みの閾値は約130dBであり、これはジェット機の離陸や至近距離でのリボルバー銃の発砲などです(Dunkle, 1982)。

図5.9 | この図は、一般的な音の大きさを示しています。(credit “planes”: modification of work by Max Pfandl; credit “crowd”: modification of work by Christian Holmér; credit: “earbuds”: modification of work by “Skinny Guy Lover_Flickr”/Flickr; credit “traffic”: modification of work by “quinntheislander_Pixabay”/Pixabay; credit “talking”: modification of work by Joi Ito; credit “leaves”: modification of work by Aurelijus Valeiša)
一般的に音の大きさに関連するのは波の振幅ですが、私たちが可聴域内の音の大きさを知覚する際には、周波数と振幅のある程度の相互作用があります。たとえば、10Hzの音波は、波の振幅に関係なく聞き取れません。一方、1000Hzの音波は、波の振幅が大きくなるにつれて、大きさの知覚の仕方が劇的に変化するでしょう。
学習へのリンク
周波数と振幅の知覚についての短い映像(http://openstax.org/l/frequency)を見て、さらに学んでください。
もちろん、異なる楽器が同じ楽譜を同じレベルの大きさで演奏しても、その響きは全く異なります。これは音の音色として知られています。音色とは、音の純粋さのことを指し、音波の周波数、振幅、タイミングの複雑な相互作用によって影響されます。
5.3 視覚
学習目標
この節が終わるまでに、あなたは次のことができるようになります:
- 視覚系の基本的な解剖学的構造を記述する
- 桿体と錐体が視覚の異なる側面にどのように貢献しているかを議論する
- 単眼手がかりと両眼手がかりがどのように奥行きの知覚に使われているかを記述する
視覚系は、私たちを取り巻く世界についての心的表象を構築します(図5.10)。これは、私たちが物理的な空間をうまく切り抜けたり、環境内の重要な個人や物体と相互作用したりする能力に貢献します。この節では、視覚系の基本的な解剖学的構造と機能の概要を提供します。加えて、私たちは色と奥行きを知覚する能力についても探求していきます。

図5.10 | 私たちの目は、私たちの周囲の世界を理解するのに役立つ感覚的な情報を取り込みます。(credit “top left”: modification of work by “rajkumar1220”/Flickr“; credit “top right”: modification of work by Thomas Leuthard; credit “middle left”: modification of work by Demietrich Baker; credit “middle right”: modification of work by “kaybee07”/Flickr; credit “bottom left”: modification of work by “Isengardt”/Flickr; credit “bottom right”: modification of work by Willem Heerbaart)
視覚系の解剖学的構造
眼球は、視覚に関わる主要な感覚器官です(図5.11)。光波は角膜を透過し、瞳孔を通って眼球に入ります。角膜は、眼球を覆う透明な膜です。角膜は、眼球の内部と外部の世界の間の障壁としての役割を果たしており、眼球に入ってきた光波の焦点を合わせることに関与しています。瞳孔は、光が通過する目の小さな開口部で、瞳孔の大きさは、光の量や感情の起伏に応じて変化します。光の量が少ない場合、瞳孔は散大、つまり拡張し、より多くの光が目に入るようになります。光の量が多い場合、瞳孔は収縮、つまり小さくなり、目に入る光の量を減らします。瞳孔の大きさは、眼球の色のついた部分である虹彩につながっている筋肉によって制御されています。
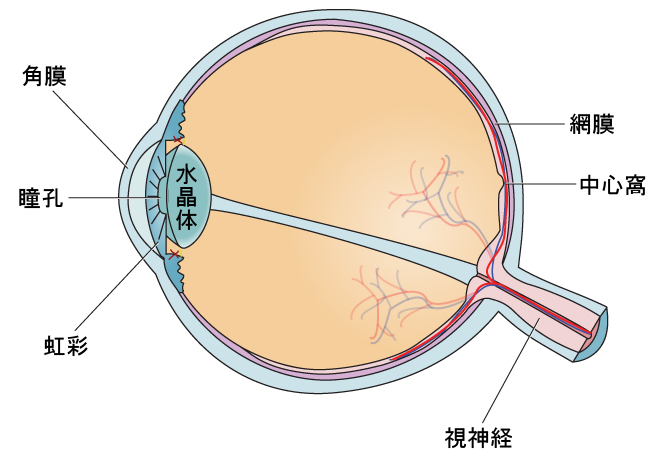
図5.11 | この図には、眼球の解剖学的構造が示されています。
瞳孔を通過した光は、水晶体を通過します。水晶体は湾曲した透明な構造体で、さらに焦点を合わせる役割を果たします。水晶体には筋肉がついており、その形を変えることで、近くや遠くの物体から反射してくる光に焦点を合わせることができます。正常な視力を持つ人の場合、水晶体は、中心窩として知られている、目の奥にある小さなくぼみに完全に像の焦点を合わせます。中心窩は、光を感知する目の内膜である網膜の一部です。中心窩には、特殊な光受容体細胞が密集しています(図5.12)。錐体として知られるこれらの光受容体細胞は、光を検知する細胞です。錐体は、明るい光の条件下で最もよく機能する特殊なタイプの光受容体です。錐体は、微小な細部に非常に敏感で、驚異的な空間分解能を与えてくれます。それらはまた、色を知覚する能力にも直接関与しています。
錐体は像が焦点を合わせられる中心窩に集中していますが、もうひとつのタイプの光受容体である桿体は、網膜の残りの部分全体に存在しています。桿体は、光の少ない条件でよく機能する特殊な光受容体です。それらには錐体のような空間分解能や色の機能はありませんが、薄暗い環境での視覚や、視野の周辺部での動きの知覚に関与しています。
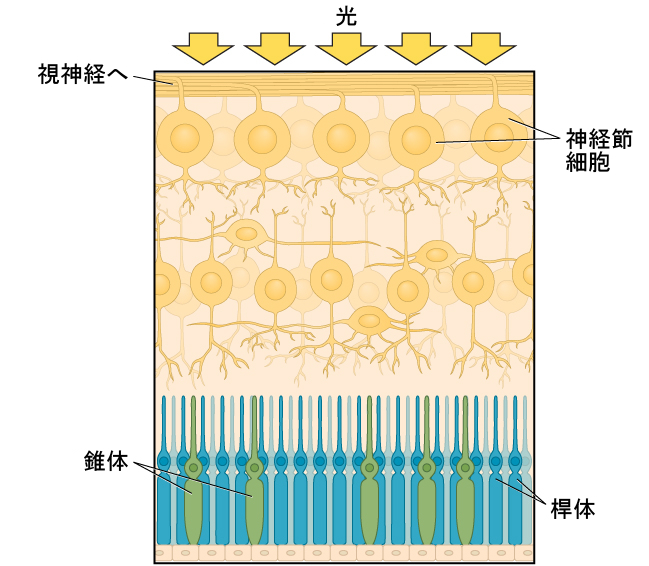
図5.12 | この図には、2種類の光受容体が示されています。錐体は緑色、桿体は青色に着色されています。
私たちは、明るく照らされた環境から薄暗い環境へ移動するときに、桿体と錐体の異なる感度を経験したことがあります。ある晴れた夏の日に超大作映画を見に行くことを想像してみてください。あなたが明るく照らされたロビーから暗い劇場の中に入ると、すぐにほとんど何も見えなくなってしまったことに気がつきます。数分後、あなたは暗さに慣れ始め、劇場の内部を見ることができるようになります。明るい環境では、主に錐体の活動があなたの視覚を支配していました。あなたが暗い環境に移ると桿体の活動が支配的になりますが、それらの段階の移行には遅れがあります。もしあなたの桿体が光を神経インパルスへと容易に、そして効果的に変換することができないと、あなたは薄暗いところでは見えにくくなります。これは、夜盲症として知られる症状です。
桿体と錐体は、(いくつかの介在ニューロンを介して)網膜神経節細胞に接続されています。網膜神経節細胞からの軸索はまとまって、目の奥から出て、視神経を形成します。視神経は、網膜からの視覚情報を脳に伝えます。視野の中には盲点と呼ばれる場所があります:盲点に小さな物体からの光が焦点を結んでも、私たちにはそれが見えません。私たちが盲点を意識的に認識することがないのには、2つの理由があります:1つ目は、それぞれの目がわずかに異なる視野の眺めを得るため、盲点が重ならないからです。2つ目は、私たちの視覚系が盲点を埋めることで、視野のその部分で発生する視覚情報に反応することはできないものの、情報が欠けていることに気づかないようにしているからです。
両目からの視神経は、脳のすぐ下にある視交叉と呼ばれる部分で合流します。図5.13が示すように、視交叉は、脳の前部の大脳皮質のすぐ下にあるX字型の構造物です。この視交叉の部分では、右側の視野からの情報(両方の眼球に由来するもの)は脳の左側に送られ、左側の視野からの情報は脳の右側に送られます。

図5.13 | この図は、脳の前部にある視交叉と、脳の後部にある後頭葉への経路を示しています。後頭葉では、視覚的な感覚が意味のある知覚へと処理されます。
脳の中に入った視覚情報は、いくつかの構造体を経由して、脳の後ろにある後頭葉に送られ、処理されます。視覚情報は、一般的に「what経路」と「where/how経路」として記述される2つの並列した経路で処理されます。「what経路」は物体の認識と識別に関係しており、「where/how経路」は空間内の位置や特定の視覚刺激との関わり方に関係します(Milner & Goodale, 2008; Ungerleider & Haxby, 1994)。たとえば、あなたが道を転がるボールを見たとき、「what経路」はその物体が何であるかを特定し、「where/how経路」は空間におけるその位置や動きを特定します。
あなたはどう考えますか?
動物を使った研究の倫理
デイヴィッド・ヒューベルとトルステン・ウィーセルは、視覚系の研究のために1981年にノーベル医学賞を授与されました。彼らは20年以上にわたって共同研究を行い、視覚的知覚の神経学についていくつもの重要な発見をしました(Hubel & Wiesel, 1959, 1962, 1963, 1970; Wiesel & Hubel, 1963)。彼らは動物、主にネコとサルを研究しました。彼らはいくつかの手法を用いましたが、中でもかなりの量の単一の単位の記録を行いました。これは、動物の脳に小さな電極を挿入し、単一の細胞がいつ活性化するかを調べるものです。彼らの多くの発見の中には、特定の方向の線には特定の脳細胞が反応すること(眼球優位性)や、それらの細胞がカラムやハイパーカラムとして知られる視覚皮質の領域にどのように配置されているかをマッピングしたことなどがありました。
彼らのいくつかの研究において、彼らは生まれたばかりの子猫の片目を縫合して閉ざし、子猫の視覚の発達を観察しました。彼らは、視覚の発達には臨界期があることを発見しました。もし子猫が片目からの入力を奪われると、視覚皮質の他の領域が、縫合して閉じられた目が通常使用する領域を埋めました。言い換えると、生まれたときに存在する神経接続は、感覚的な入力を奪われると失われる可能性があるということです。
研究のために子猫の目を縫って閉じることについて、あなたはどう考えますか?多くの動物愛護家にとって、これは残忍で、虐待的で、非倫理的であると思われるでしょう。もし、ある症状を持って生まれた赤ん坊や子供が、目が見えなくなるのではなく、正常な視覚を発達させられるのを助けるような研究ができるとしたらあなたはどうしますか?あなたはそのような研究をしたいと望みますか?猫に多少の危害を加えることになったとしても、あなたはその研究を行うでしょうか?もしあなたがそのような子供の親であったとしたら、同じように考えるでしょうか?もし、あなたが動物保護施設で働いていたらどうでしょうか?
他のほぼすべての先進国と同様に、アメリカでも動物を使った医学実験は、(十分な科学的正当性があると仮定して)ほとんど制限なく認められています。既存の法律の目的は、そのような実験を禁止することではなく、実験室での動物の人道的な扱いや収容に関する基準を設けることによって、不必要な動物の苦痛を抑えることにあります。
イェール大学にある生命倫理学学際センターの所長スティーブン・レイサムが説明しているように(Latham, 2012)、動物実験に対する法律や規制のあり得るアプローチは、政府による強力な規制とすべての実験の監視から、研究者の倫理観に依拠した自主規制のアプローチまで、連続性の中でさまざまに異なります。イギリスは最も顕著な規制スキームを持っている一方で、日本は自主規制のアプローチを採用しています。米国のアプローチはその中間に位置し、2つのアプローチが徐々に混ざり合った結果となっています。
医学研究が有益かつ重要な実践であることに疑問の余地はありません。問題は、最も信頼性の高い結果を得るために、動物を使用することが必要なのか、あるいは最良の実践なのかどうかということです。代替手段としては、患者と薬剤のデータベース、仮想薬剤試験、コンピュータモデルとシミュレーション、磁気共鳴画像法やコンピュータ断層撮影法などの非侵襲的画像技術を使うことが含まれます(“Animals in Science/Alternatives,” n.d.)。また、マイクロドージング[超微量投与]などの他の技術では、人間を実験動物としてではなく、試験結果の正確性や信頼性を向上させるための手段として用います。また、人間の細胞や組織の培養、幹細胞、および遺伝子検査法に基づくインビトロ法もますます利用可能になっています。
今日、地方レベルでは、動物を使用し、連邦政府から資金援助を受けている施設は、NIHのガイドラインが遵守されていることを確認する機関内動物ケア・使用委員会(IACUC)を設置しなければなりません。IACUCには、研究者、管理者、獣医師、そしてその機関とは関係のない少なくとも1人の人物、すなわち関心のある市民が含まれていなければなりません。この委員会は、研究室や手順の監査も行います。
色と奥行きの知覚
私たちは世界を白黒で見ているわけではなく、二次元(2-D)、すなわち平面(高さと幅だけで、奥行きがない)で見ているわけでもありません。ここでは、色覚がどのように働くのかと、私たちがどのようにして三次元(高さ・幅・奥行き)を知覚するのかについて見ていきましょう。
色覚
通常の視力の人は、色覚をつかさどる3つの異なる種類の錐体を有しています。それぞれの種類の錐体は、わずかに異なる波長の光に対して最大の感度を持っています。図5.14に示される色覚の三色理論によると、スペクトル内のすべての色は、赤色、緑色、青色を組み合わせることによって生成されます。3種類の錐体はそれぞれ、それらの色のうちの1つを受容します。
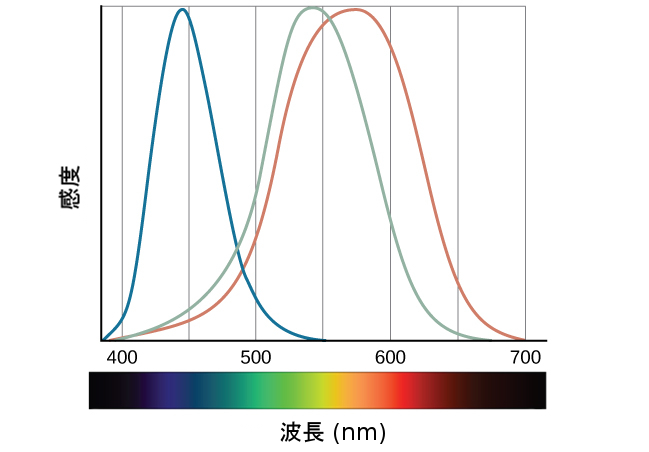
図5.14 | この図は、通常の視力の人に見られる3種類の錐体の感度の違いを示しています。(credit: modification of work by Vanessa Ezekowitz)
概念をつなげてみよう
色覚異常:ある個人的なストーリー
数年前、私は人前に出るための服を着て、7歳の娘が座っているキッチンに入っていきました。彼女は私を見上げて、最も厳しい声音で「それは着たらダメ」と言いました。私が「どうしてダメなんだい?」と尋ねると、娘は私の服の色が合っていないことを教えてくれました。彼女は私がシャツやズボン、ネクタイの色を揃えるのが下手だとよく文句を言っていましたが、今回は特に深刻そうに聞こえました。シングルファーザーの私は、家の中では他に尋ねる人もいないので、近くのコンビニエンスストアに車で行き、店員に服の色が合っているかどうかを尋ねました。すると彼女は、私のズボンは鮮やかな緑色、シャツは赤みがかったオレンジ色、ネクタイは茶色だと言うのです。彼女は私をいぶかしげに見て、「あなたの服が合っているわけがない」と言いました。それから数日の間、私は同僚や友人に自分の服が合っているかどうかを尋ねるようになりました。同僚たちから、ただ単に私が「本当に個性的なスタイル」だと思っていたと言われること数日の後、私は眼科医に予約を入れて検査を受けました(図5.15)。その時になって、私は自分が色覚異常であることがわかりました。私はほとんどの緑色、茶色、赤色の区別がつきません。幸いなことに、知らず知らずのうちに服装がひどいことになっていること以外は、私の色覚異常が日常生活に支障をきたすことはほとんどありません。

図5.15 | 石原検査は、色や大きさの異なるドットからなる円の中に現れる数字を識別できるかどうかを検査することによって色の知覚を評価します。
色覚異常の中には、まれな形態もあります。グレースケール(黒と白の濃淡のみ)で見ている人は極度にまれであり、そのような人は桿体しか持っていません。これは、その人たちは視力が非常に低く、あまりよく見ることができないことを意味します。最も一般的なX連鎖遺伝性異常は、赤-緑色覚異常です(Birch, 2012)。ヨーロッパのコーカサス出自の男性の約8%、アジア系男性の約5%、アフリカ系男性の約4%、米国先住民男性、オーストラリア系男性、ポリネシア系男性の約2%未満が赤-緑色覚異常を有しています(Birch, 2012)。一方、ヨーロッパのコーカサス出自の女性では、赤-緑色覚異常は約0.4%に過ぎません(Birch, 2012)。
色覚の三色理論が唯一の理論というわけではありません。もう1つの色覚の主要な理論は、反対-処理理論として知られています。この理論によると、色は黒色-白色、黄色-青色、緑色-赤色という反対のペアでコード化されます。基本的な考え方としては、視覚系の一部の細胞は、反対色の一方の色によって励起され、もう一方の色によって抑制されるというものです。つまり、緑色に関連する波長によって励起される細胞は、赤色に関連する波長によって抑制され、その逆もまた然りです。反対処理の意味合いの1つは、私たちが緑色がかった赤色や黄色がかった青色を色として経験しないということです。もう1つの意味合いは、これが陰性の残像の経験につながるということです。残像とは、刺激を取り除いた後も視覚的な感覚が継続することを言います。たとえば、あなたが太陽を少し見てから目を離すと、刺激(太陽)がなくなったにもかかわらず、光の点をまだ感じることがあります。刺激に色が含まれている場合、反対-処理理論で特定された色の組み合わせによって、陰性の残像がもたらされます。あなたはこの概念を、図5.16の旗を使って試してみることができます。

図5.16 | 白い点を30~60秒見つめた後、白紙に目を移してみてください。あなたには何が見えますか?これは陰性の残像として知られているものであり、色覚の反対-処理理論の実証的な裏付けを与えてくれます。
しかし、この2つの理論(色覚の三色理論と反対-処理理論)は、相互に排他的ではありません。研究の結果、これらは神経系の異なるレベルに適用されるものだということが示されています。網膜での視覚処理では、三色理論が適用されます:錐体は、赤色、青色、緑色を表す3つの異なる波長に反応します。しかし、信号が網膜を通過して脳に到達すると、細胞は反対-処理理論に合致した方法で反応します(Land, 1959; Kaiser, 1997)。
学習へのリンク
色の知覚についてのビデオ(http://openstax.org/l/colorvision)を見て、さらに学んでください。
奥行き知覚
私たちが三次元(3-D)空間において空間的な関係を知覚する能力は、奥行き知覚として知られています。奥行き知覚では、私たちは物体が他の物体の前、後ろ、上、下、または横にあると表現することができます。
私たちの世界は三次元であり、私たちによるこの世界の心的表象が三次元の特性を持つのは納得がいきます。私たちは、視覚的な光景の中で、さまざまな手がかりを使って奥行きの感覚を確立します。そのうちの一部は、両眼手がかりです。これは、それらが両方の目の使用に依拠していることを意味します。両眼の奥行きの手がかりの一例は、両眼視差、すなわち、私たちの両眼のそれぞれがわずかに異なる世界の見え方を受け取ることです。このわずかに異なる見え方を体験するには、次のような簡単な実験をやってみてください:あなたの腕をいっぱいに伸ばし、一本の指を伸ばして、その指に焦点を合わせます。そして、頭を動かさずに左目を閉じ、次に頭を動かさずに左目を開いて右目を閉じます。2つの目で交互に見ると、指が移動しているように見えることに気づくでしょう。これは、それぞれの目がわずかに異なる指の見え方をしているためです。
3-D映画も同じ原理で機能しています:あなたが特殊なメガネをかけることによって、スクリーンに映し出される2つのわずかに異なる画像を、左目と右目で別々に見ることができます。脳がこれらの画像を処理することで、飛び跳ねる動物や走る人がまっすぐ自分の方に向かってきているように錯覚します。
私たちは3-D世界において奥行きを経験するために両眼手がかりに頼っていますが、私たちはまた、二次元の配列において奥行きを知覚することもできます。あなたがこれまでに見たことのあるすべての絵画や写真を思い出してみてください。一般的には、視覚刺激が二次元であっても、あなたはそれらの画像の中において奥行きを受け取ります。私たちがこれを行うとき、私たちはいくつかの単眼手がかり、つまり片方の目のみを必要とする手がかりに頼っています。もしあなたが、片目では奥行きが見えないと思っているならば、片目のみを使って歩いているときでも物にぶつかることがない、ということに留意してみてください。そして、実際に、私たちは両眼手がかりよりも多くの単眼手がかりを有しています。
単眼手がかりの一例としては、線遠近法として知られるものがあります。線遠近法とは、私たちが画像の中で収束するように見える2本の平行線を見たときに、奥行きを知覚する事実のことを指します(図5.17)。他の単眼の奥行きの手がかりとしては、介在、物体の部分的な重なり、および地平線に対する画像の相対的な大きさや近さがあります。

図5.17 | 私たちは、線遠近法(道路が遠くに向かって狭くなるにつれて平行線が収束していくようなもの)のような単眼手がかりを使って、このような二次元の図の中で奥行きを知覚します。(credit: Marc Dalmulder)
深く掘り下げてみよう
立体視異常
ブルース・ブリッジマンは、生まれつき極端な斜視で、両眼の奥行きの手がかりに反応できない立体視異常を持っていました。彼は単眼の奥行きの手がかりに頼ることが多かったのですが、自分の周りの世界の3-Dの性質を真に理解することはありませんでした。2012年のある夜、ブルースが妻と一緒に映画を観に行ったときに、すべてが変わりました。
2人が観に行った映画は3-Dで撮影されていました。ブルースはお金の無駄だと思いながらも、チケット購入時に3-Dメガネの代金を支払いました。映画が始まるやいなや、ブルースはメガネをかけて全く新しい体験をしました。彼は生まれて初めて、自分を取り巻く世界の本当の奥行きを理解したのです。驚くべきことに、彼の奥行きを知覚する能力は映画館の外でも持続しました。
神経系には、両眼の奥行きの手がかりに反応する細胞があります。通常、これらの細胞が持続するためには、発達の初期段階で活性化される必要があります。そのため、ブルースの事例(および彼と似たような他の事例)に詳しい専門家は、ブルースが発達のある時点で、少なくとも一瞬でも両眼視を経験したに違いないと推測しています。それは、両眼手がかりに対応する視覚系の細胞が生き残るためには十分なものでした。現在の謎は、なぜブルースがこれらの細胞を活性化させるのに70年近くかかったのかということです(Peck, 2012)。
5.4 聴覚
学習目標
この節が終わるまでに、あなたは次のことができるようになります:
- 聴覚系の基本的な解剖学的構造と機能を記述する
- 私たちがどのようにして音の高さをコード化し、知覚するかを説明する
- 私たちがどのようにして音を定位するかを議論する
私たちの聴覚系は、圧力波を意味のある音へと変換します。このことは、私たちが自然の音を聞いたり、音楽の美しさを堪能したり、話し言葉を通じてお互いにコミュニケーションをとる能力につながっています。この節では、聴覚系の基本的な解剖学的構造と機能についての概要を提供します。そこには、感覚刺激がどのようにして神経インパルスに変換されるのか、その情報は脳のどこで処理されるのか、私たちがどのようにして音の高さを知覚するのか、私たちは音がどこから聞こえてくるのかをどのようにして知るのか、などが含まれます。
聴覚系の解剖学的構造
耳は複数の区域に分けることができます。外耳には、私たちの頭部から突き出た目に見える耳の部分である耳介、耳道、および鼓膜があります。中耳には、耳小骨として知られる3つの小さな骨があります。これらの骨は、槌骨、砧骨、あぶみ骨と名前が付けられています。内耳には、バランスや運動(前庭感覚)に関わる半規管と、蝸牛とがあります。蝸牛は、液体で満たされたカタツムリのような形をした構造で、聴覚系の感覚受容体細胞(有毛細胞)を収容しています(図5.18)。
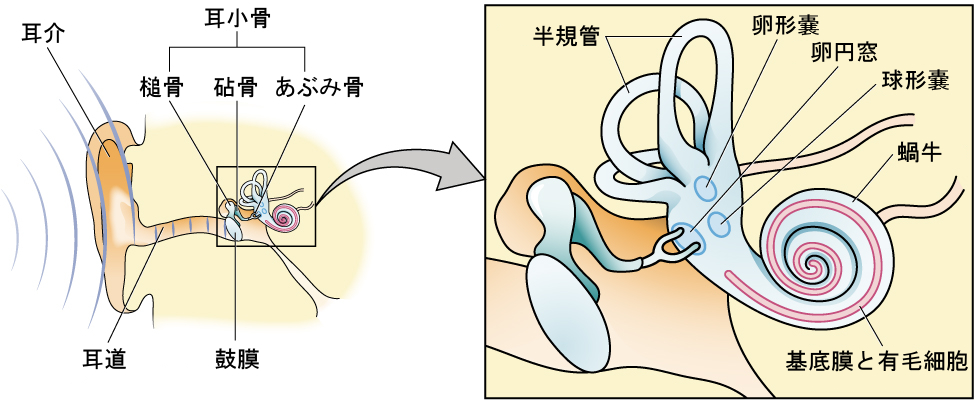
図5.18 | 耳は、外耳(耳介と鼓膜)、中耳(3つの耳小骨:槌骨、砧骨、あぶみ骨)、内耳(蝸牛と基底膜)に分かれています。
音波は耳道を伝わって鼓膜にぶつかり、鼓膜を振動させます。この振動により、3つの耳小骨が動きます。耳小骨が動くと、あぶみ骨が卵円窓として知られる蝸牛の薄い膜に押し付けられます。あぶみ骨が卵円窓を押すと、蝸牛内の液体が動き出し、基底膜に埋め込まれた内耳の聴覚受容体細胞である有毛細胞が刺激されます。基底膜は蝸牛の中にある薄い一片の組織です。
有毛細胞の活性化は機械的なプロセスです:有毛細胞への刺激が最終的に有毛細胞の活性化につながります。有毛細胞が活性化されると、神経インパルスが発生し、これが聴神経を通って脳に到達します。聴覚情報は、下丘、視床の内側膝状核、そして最後に脳の側頭葉の聴覚皮質に送られ、処理されます。また、視覚系と同様に、聴覚の認識や定位についての情報は並列の流れで処理されていることを示唆する証拠もあります(Rauschecker & Tian, 2000; Renier et al., 2009)。
音の高さの知覚
音波の異なる周波数は、その音の高さに対する私たちの知覚の違いに関連付けられます。低い周波数の音は低音であり、高い周波数の音は高音です。聴覚系はどのようにしてさまざまな音の高さを区別しているのでしょうか?
音の高さの知覚を説明するために、いくつかの理論が提唱されています。私たちはここでは、それらのうちの2つについて議論します:それは時間理論と場所理論です。音の高さの知覚についての時間理論は、周波数が感覚ニューロンの活動レベルによってコード化されると主張します。これは、所与の有毛細胞が音波の周波数に関連した活動電位を発火することを意味するでしょう。これは非常に直感的な説明ではありますが、私たちが検出する周波数は20~20000Hzと非常に幅広いため、有毛細胞が発火する活動電位の周波数ではすべての範囲の周波数を説明することができません。活動電位に関与する神経膜上のナトリウムチャネルに関する特性のために、細胞がそれ以上速く発火できないようなポイントがあります(Shamma, 2001)。
音の高さの知覚についての場所理論は、基底膜の異なる部分が異なる周波数の音に対して感受性があると示唆しています。より具体的には、基底膜の基部は高い周波数に最もよく反応し、基底膜の先端部は低い周波数に最もよく反応します。したがって、基底膜の基部にある有毛細胞は高音域の受容体、基底膜の先端部にある有毛細胞は低音域の受容体と分類されることになるでしょう(Shamma, 2001)。
実際には、どちらの理論も音の高さの知覚の異なる側面を説明しています。約4000Hzまでの周波数では、活動電位の速度と場所の両方が音の高さの知覚に寄与していることは明らかです。しかしながら、はるかに高い周波数の音は、場所の手がかりを使ってのみコード化されています(Shamma, 2001)。
音の定位
私たちの環境の中で音の位置を定位する能力は、聴覚の重要な部分です。音の定位は、私たちが視野の中で奥行きを知覚する方法に似ていると考えられます。奥行きについての情報を提供する単眼手がかりおよび両眼手がかりと同様に、聴覚系は単耳手がかり(モノラル:1つの耳)および両耳手がかり(バイノーラル:2つの耳)を使用して音を定位します。
音源と体の位置関係に応じて、それぞれの耳介は入ってくる音波と異なった形で相互作用します。この相互作用が単耳手がかりを提供し、それは私たちの上下や前後で生じた音の位置を特定するのに役立ちます。真上、真下、真正面、または真後ろからやって来た音を両耳で受け取った音波は同じになるので、単耳手がかりは不可欠なものです(Grothe, Pecka, & McAlpine, 2010)。
一方、両耳手がかりは、両耳の鼓膜の振動パターンの違いに基づくことにより、水平軸に沿った音の位置の情報を与えてくれます。もし音が中心から外れた位置から来た場合、両耳間レベル差と両耳間タイミング差という2種類の両耳手がかりが発生します。両耳間レベル差とは、あなたの体の右側からやって来た音は、頭部を通過する際に音波が減衰するため、右耳では左耳よりも強く聞こえるという事実のことを指します。両耳間タイミング差とは、ある音波がそれぞれの耳に到達する時間のわずかな違いのことを指します(図5.19)。脳の特定の領域がこの差を監視して、音が水平軸上のどこから発生したかを構築します(Grothe et al., 2010)。
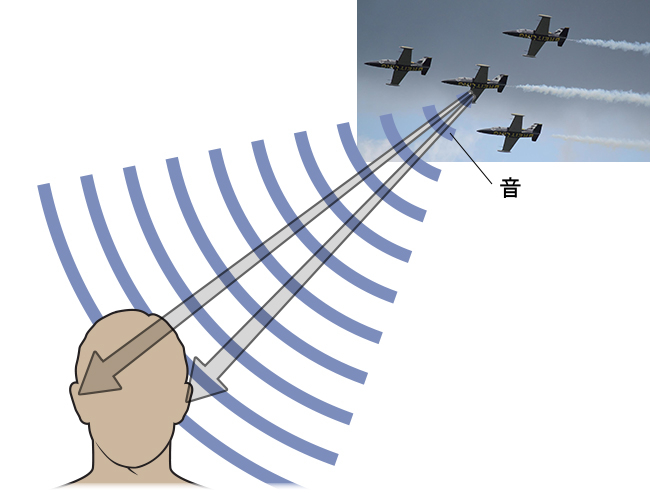
図5.19 | 音の定位には、単耳手がかりと両耳手がかりの両方が使われます。(credit “plane”: modification of work by Max Pfandl)
聴覚の喪失
聴覚障害とは、部分的または完全に聞くことのできない状態のことです。一部の人は生まれつき耳が聞こえず、これは先天性難聴と呼ばれています。他の人は伝音性難聴であり、これは音のエネルギーを蝸牛に届けるのに問題があることに起因します。伝音性難聴の原因には、外耳道の閉塞、鼓膜の穴、耳小骨の問題、鼓膜と蝸牛の間の空間の液体などがあります。他の人の一群は、最も一般的な難聴である感音性難聴です。感音性難聴は、加齢、頭部外傷や音響外傷、感染症や病気(麻疹やおたふく風邪など)、薬剤、騒音曝露などの環境的影響(図5.20に示される騒音性難聴)、腫瘍、毒素(特定の溶剤や金属に含まれるものなど)など、多くの要因によって引き起こされます。

図5.20 | 感音性難聴につながり得る環境要因には、大音量の音楽や建設機械に定期的にさらされることが含まれます。(a)音楽演奏者や(b)建設作業員はこの種の難聴のリスクがあります。(credit a: modification of work by “GillyBerlin_Flickr”/Flickr; credit b: modification of work by Nick Allen)
音波の刺激が鼓膜から耳小骨を経て蝸牛の卵円窓に伝達されるという機械的な性質を考慮すると、ある程度の難聴は避けられません。伝音性難聴では、聴覚の問題は、鼓膜の振動および/または耳小骨の動きがうまくいかないことに関連しています。このような問題は、入ってくる音波を増幅して鼓膜の振動や耳小骨の動きを起こりやすくする補聴器などの機器によってしばしば対処されます。
蝸牛から脳への神経信号の伝達がうまくいかないことに関連する聴覚の問題は、感音性難聴と呼ばれます。感音性難聴の原因となる病気の1つにメニエール病があります。あまりよく理解されていませんが、メニエール病では内耳の構造が変性し、聴覚の喪失、耳鳴り(常に音が鳴り響いていたり雑音がしたりする状態)、めまい(回転しているような感覚)、および内耳内の圧力の上昇につながることがあります(Semaan & Megerian, 2011)。この種の難聴は、補聴器で治療することはできませんが、一部の人は、治療の選択肢として人工内耳を使用する対象者となり得ます。人工内耳は、マイク、音声処理装置、および電極アレイで構成される電子機器です。この装置は、やって来る音の情報を受け取り、聴神経を直接刺激して脳に情報を伝達します。
学習へのリンク
人工内耳の手術についてのビデオ(http://openstax.org/l/cochlear)を見て、さらに学んでください。
あなたはどう考えますか?
聾文化
アメリカや世界の他の場所では、聴覚障害者は自分たち自身の言語、学校、および習慣を持っています。これは聾文化と呼ばれます。アメリカでは、聾者はしばしばアメリカ手話(ASL)を使ってコミュニケーションをとります。ASLには口頭の要素がなく、視覚的な手話やジェスチャーだけで成り立っています。主なコミュニケーションの様式は手話です。聾文化の価値観のひとつは、聾者の子供たちに対して言葉を話したり、唇を読んだり、人工内耳の手術をしたりすることを教えるのではなく、手話を使うことのような伝統を続けることです。
子供が耳が聞こえないと診断されたとき、親は難しい決断を迫られます。その子を主流の学校に入学させ、言葉の発音の仕方や唇の読み方を教えるべきでしょうか?それとも、その子を聴覚障害児のための学校に通わせ、ASLを学び、聾文化に触れる機会を増やすべきでしょうか?親が聾者であるか否かによって、これらの決定に対する親のアプローチの仕方に違いがあると考えますか?
5.5 その他の感覚
学習目標
この節が終わるまでに、あなたは次のことができるようになります:
- 化学感覚の基本的な機能を記述する
- 体性感覚系、侵害受容感覚系、および熱受容感覚系の基本的な機能を説明する
- 前庭感覚系、固有受容感覚系、および運動感覚系の基本的な機能を記述する
視覚と聴覚は、長年にわたって研究者から非常に多くの注目を集めてきました。これらの感覚系がどのように機能するかについては、まだ多くの学ぶべきことが残っていますが、私たちは他の感覚の様式に比べればそれらのことをはるかによく理解しています。この節では、私たちは、化学感覚(味と匂い)と身体感覚(接触、温度、痛み、バランス、体勢)について探求していきます。
化学感覚
味(味覚)と匂い(嗅覚)は化学感覚と呼ばれています。なぜなら、どちらも私たちが口にする食べ物や呼吸する空気に含まれる分子に反応する感覚受容体を持っているからです。化学感覚の間では、顕著な相互作用があります。たとえば、私たちが食べ物の風味を表現するときには、私たちは実際にはその食べ物の味覚と嗅覚の両方の特性を組み合わせて言及します。
味(味覚)
あなたは、味覚には4つの基本的なグループがあることを小学校の頃から学んでいます:それは、甘味、塩味、酸味、苦味です。しかしながら、研究によると、私たちには少なくとも6つの味覚グループがあることがわかっています。うま味は私たちの5番目の味です。うま味は、実際には日本語の単語であり、大雑把には「おいしい」と翻訳することができます。うま味は、グルタミン酸ナトリウムの味と関連しています(Kinnamon & Vandenbeuch, 2009)。また、私たちが食べ物の脂肪分に対する味覚を持っていることを示唆する実験的証拠も増えています(Mizushige, Inoue, & Fushiki, 2007)。
私たちが摂取した食べ物や飲み物の分子は、唾液に溶け込み、舌や口の中、喉にある味覚受容体と相互作用します。味蕾は、味蕾の中心孔に突き出た毛状の伸長部分を持つ味受容体細胞の集まりで形成されています(図5.21)。味蕾の寿命は10日から2週間なので、舌をやけどすることによっていくつかを破壊しても長期的な影響はなく、それらはすぐに生えてきます。味の分子は、この伸長部分にある受容体に結合し、感覚細胞内で化学変化を起こします。その結果、神経インパルスが受容体の位置に応じて異なる神経を介して脳に伝達されます。味覚情報は、延髄、視床、および大脳辺縁系へと、そして前頭葉と側頭葉の重なりの下に押し込まれている味覚皮質へと伝達されます(Maffei, Haley, & Fontanini, 2012; Roper, 2013)。
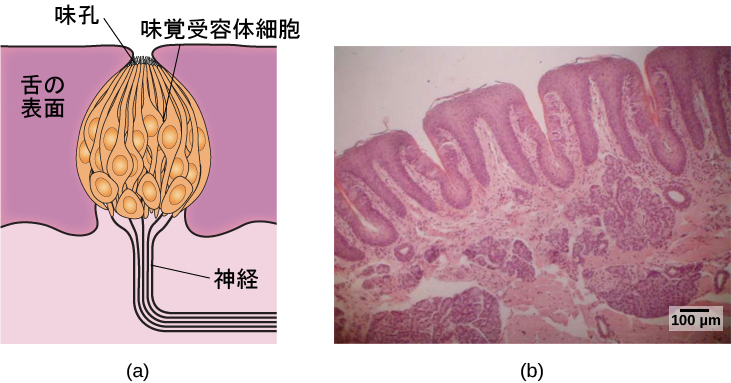
図5.21 | (a)味蕾は、神経に情報を伝達する個々の味覚受容体細胞が多数集まって構成されています。(b)この顕微鏡写真は、舌の表面をクローズアップした図を示しています。(credit a: modification of work by Jonas Töle; credit b: scale-bar data from Matt Russell)
匂い(嗅覚)
嗅覚受容体細胞は、鼻の上部にある粘膜の中に位置しています。この受容体から出ている小さな毛のような伸長部分が、粘液に溶け込んでいる匂いの分子と、その伸長部分にある化学受容体との相互作用の場となります(図5.22)。匂いの分子が受容体に結合すると、細胞内で化学変化が起こり、信号が嗅球(前頭葉の先端にある球状の構造で、嗅神経が始まる場所)に送られます。嗅球からの情報は、大脳辺縁系の領域や、味覚皮質のすぐ近くに位置する一次嗅覚皮質に送られます(Lodovichi & Belluscio, 2012; Spors et al., 2013)。
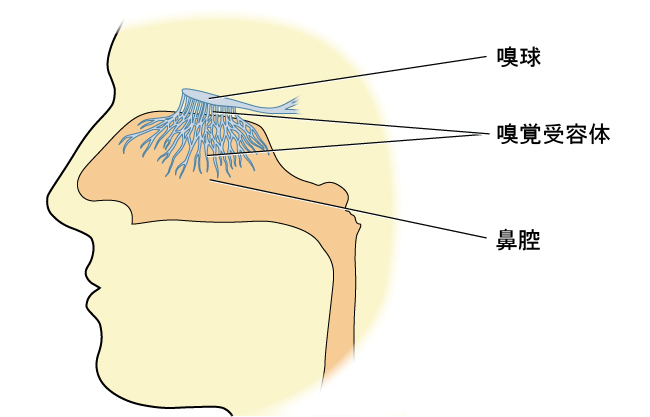
図5.22 | 嗅覚受容体は、嗅球から鼻腔の粘膜内に伸びる毛のような部分です。
異なる種の嗅覚系の感度には、非常に大きなばらつきがあります。私たちは、犬が人間よりもはるかに優れた嗅覚系を持っているとしばしば考えており、実際に、犬はその鼻によっていくつかの驚くべきことを行うことができます。犬は血糖値の危険な低下やがんの腫瘍を「嗅ぐ」ことができるという証拠もあります(Wells, 2010)。犬の並外れた嗅覚能力は、嗅覚受容体のための機能的な遺伝子の数が、人間や他の霊長類では400個未満であるのに対し、犬では800~1200個と多いことによるのかもしれません(Niimura & Nei, 2007)。
多くの生物種は、他の個体から送られてくるフェロモンとして知られる化学的メッセージに反応します(Wysocki & Preti, 2004)。フェロモンによるコミュニケーションには、しばしば潜在的な交尾相手の生殖状態についての情報を提供することが含まれます。たとえば、雌のラットが交尾の用意ができたときには、フェロモンシグナルを分泌して、近くにいる雄のラットの注意を引きます。フェロモンの活性化は、実際には雄のラットの性行動を引き出すための重要な要素です(Furlow, 1996, 2012; Purvis & Haynes, 1972; Sachs, 1997)。また、人間のフェロモンについても多くの研究(および論争)が行われています(Comfort, 1971; Russell, 1976; Wolfgang-Kimball, 1992; Weller, 1998)。
触覚、熱受容感覚、侵害受容感覚
皮膚には、接触に関連するさまざまな刺激に反応する多くの受容体が分布しています(図5.23)。これらの受容体には、マイスナー小体、パチニ小体、メルケル盤、ルフィニ小体が含まれます。マイスナー小体は圧力や低周波数の振動に反応し、パチニ小体は一時的な圧力や高周波数の振動を感知します。メルケル盤は軽い圧力に反応し、ルフィニ小体は伸縮を感知します(Abraira & Ginty, 2013)。
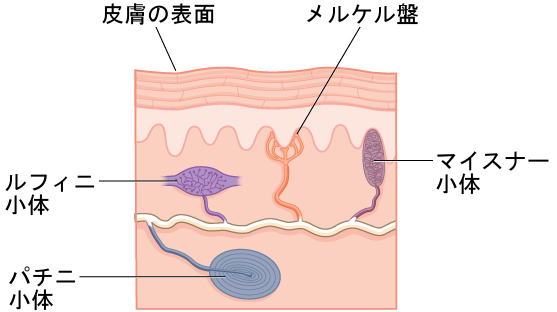
図5.23 | 皮膚には多くの種類の感覚受容体が存在し、それぞれが特定の接触に関連した刺激に反応します。
皮膚にある受容体に加えて、感覚の機能に役立つ自由神経終末が多数存在します。これらの神経終末は、さまざまな異なる種類の接触に関連した刺激に反応し、熱受容(温度の知覚)と侵害受容(潜在的な危害およびおそらく痛みを示す信号)の両方の感覚受容体として機能します(Garland, 2012; Petho & Reeh, 2012; Spray, 1986)。受容体と自由神経終末から集められた感覚情報は、脊髄を伝わり、延髄、視床、そして最終的には頭頂葉の中心後回に位置する体性感覚皮質に伝達されます。
痛みの知覚
痛みは、身体的および心理的な要素を含む不快な経験です。痛みを感じることは、私たちに対して怪我をしたことを認識させ、その怪我の原因から逃れようとする動機となるため、非常に適応的なものです。さらに、痛みを感じることで、私たちは怪我をした体の部分に優しく接することになるので、さらに怪我をする可能性が低くなります。
一般的に言って、痛みはその性質において神経障害性のものか、あるいは炎症性のものと見なされます。あるタイプの組織損傷を知らせる痛みは、炎症性疼痛として知られています。いくつかの状況では、末梢神経系または中枢神経系のニューロンの損傷によって痛みが引き起こされます。その結果、脳に送られる痛みの信号が誇張されます。このような種類の痛みは神経障害性疼痛として知られています。痛みを和らげるための複数の治療の選択肢は、リラクセーション療法から、鎮痛剤の使用、脳深部刺激法に至るまで、多岐にわたっています。ある個人にとって最も効果的な治療法は、痛みの程度や持続性、医学的/心理的条件などを含むさまざまな考慮事項に基づいて、異なったものとなります。
一部の人は、生まれつき痛みを感じる能力を持ち合わせていません。この非常に稀な遺伝性疾患は、先天性無痛覚症として知られています。先天性無痛覚症の人は、温度や圧力の違いを感知することはできますが、痛みを経験することができません。その結果、彼らはしばしば大きな傷を負うことがあります。幼い子供たちは、自分で何度も噛んだために、口や舌に深刻な傷を負っています。驚くことではありませんが、この障害を持つ人は、その傷や傷ついた部位の二次感染のために、平均余命が非常に短くなります(U.S. National Library of Medicine, 2013)。
学習へのリンク
先天性無痛覚症についてのビデオ(http://openstax.org/l/congenital)を見て、さらに学んでください。
前庭感覚、固有受容感覚、運動感覚
前庭感覚は、私たちがバランスと体の姿勢を維持する能力に貢献しています。図5.24が示すように、この系の主要な感覚器官(卵形嚢、球形嚢、三半規管)は、内耳の蝸牛の隣に位置しています。前庭器官は液体で満たされており、聴覚系に見られるものと似た、頭の動きや重力に反応する有毛細胞があります。この有毛細胞が刺激を受けると、それらは前庭神経を介して脳に信号を送ります。私たちは通常の状況下では、前庭系の感覚情報を意識的に認識することはありませんが、私たちが乗り物酔いおよび/または内耳の感染症に関連するめまいを経験するときには、その重要性が明らかになります(Khan & Chang, 2013)。
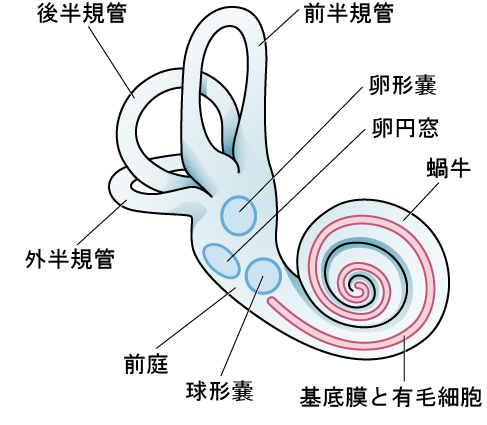
図5.24 | 前庭系の主要な感覚器官は、内耳の蝸牛の隣に位置しています。これらの器官には、卵形嚢、球形嚢、および三半規管(後半規管、前半規管、外半規管)が含まれます。
前庭系は、バランスを維持することに加えて、体の位置の変化を補うために体のさまざまな部分を動かす運動と反射を制御するのに重要な情報を収集します。そのため、前庭系が提供する情報を利用して、固有受容感覚(体の位置の知覚)と運動感覚(空間における身体の動きの知覚)が相互作用します。
また、これらの感覚系は、筋肉、関節、皮膚、および腱の伸縮や緊張に反応する受容体からも情報を収集します(Lackner & DiZio, 2005; Proske, 2006; Proske & Gandevia, 2012)。固有受容感覚と運動感覚の情報は、脊柱を経由して脳に伝えられます。小脳に加えていくつかの皮質領域が、固有受容感覚系と運動感覚系の感覚器官から情報を受け取り、情報を送ります。
5.6 知覚についてのゲシュタルト原理
学習目標
この節が終わるまでに、あなたは次のことができるようになります:
- 図と地の関係性を説明する
- グループ分けのゲシュタルト原理を定義する
- 知覚的構えが個人の特性や精神状態によってどのように影響されるかを記述する
20世紀初頭、マックス・ヴェルトハイマーは、人が急速に点滅する静止画像において動きを知覚することを実証した論文を発表しました(その洞察は、彼が子供のおもちゃのタキストスコープを使ったときに思いついたものでした)。ヴェルトハイマーは、後にパートナーとなる助手のヴォルフガング・ケーラーとクルト・コフカとともに、知覚は単に感覚刺激を組み合わせたもの以上のことが関わっていると考えました。この考えは、心理学の分野でゲシュタルト心理学として知られる新しい動きへとつながりました。ゲシュタルトという言葉は、字義通りには「形態」や「パターン」を意味しますが、その使用は「全体は部分の総和とは異なる」という考え方を反映しています。言い換えれば、脳は利用可能な感覚入力の単純な合計以上の知覚を作り出しており、それは予測可能な方法で行われます。ゲシュタルト心理学者は、この予測可能な方法を、私たちが感覚情報を整理するための原理へと言い換えました。その結果、ゲシュタルト心理学は感覚と知覚の分野で非常に大きな影響力を持っています(Rock & Palmer, 1990)。
ゲシュタルト原理の1つに、図と地の関係性があります。この原理によると、私たちは視覚世界を図と地に分ける傾向があります。図とは視野の焦点となる物体や人物のことで、地とは背景のことです。図5.25が示すように、何を図と認識し、何を地と認識するかによって、私たちの知覚は大きく変わります。おそらく、感覚情報を解釈する私たちの能力は、何らかの特定の状況において私たちが何を図とし、何を地とするかに基づくものですが、この仮定には疑問が投げかけられています(Peterson & Gibson, 1994; Vecera & O’Reilly, 1998)。

図5.25 | 図と地の関係性の概念は、この画像が花瓶としても一対の顔としても知覚することができる理由を説明してくれます。
感覚的な刺激を意味のある知覚に整理するためのもう1つのゲシュタルト原理は、近接です。この原理は、図5.26が示すように、お互いに近くにあるものは一緒にグループ化される傾向があるということを主張しています。

図5.26 | 近接のゲシュタルト原理は、(a)左側では丸の1つのかたまりが見え、(b)右側では3つの列が見えることを示唆しています。
私たちが何かをどのように読むかということも、近接の概念を示してくれます。たとえば、「We read this sentence like this, notl iket hiso rt hat.」(私たちはこの文章をこのように読み、こ のように はよ みませ ん)。私たちは、文字と文字の間にスペースがないことによってある単語の文字をグループ化し、それぞれの単語の間にスペースがあることによって単語を認識します。さらにいくつかの例を紹介します:「Cany oum akes enseo ft hiss entence? What doth es e wor dsmea n?」(あな たはこ のぶん しょうのい みがわかりま すか?これ らのこと ばはなに をい みしてい ますか?)
私たちはまた、類同の原理を利用して、視野の中のものをグループ化することもあります。この原理によると、似ているものは一緒にグループ化される傾向があります(図5.27)。たとえば、フットボールの試合を見ているとき、私たちはユニフォームの色に基づいて個人をグループ化する傾向があります。攻撃側のドライブを見るとき、この特徴でグループ化するだけで、2つのチームを感得することができます。

図5.27 | この丸の配列を見ると、私たちは色の列が交互に並んでいるように知覚します。私たちは、類同の原則に従って、これらの丸をグループ化しています。
さらなる2つのゲシュタルト原理に、連続(あるいは、よい連続)の法則と閉合の法則があります。連続の法則は、私たちがギザギザした折れ線よりも、連続した滑らかな流れのある線を知覚しやすいということを示唆します(図5.28)。閉合の原理は、私たちが知覚を一連の部品としてではなく、完全な物体として整理するということを述べています(図5.29)。

図5.28 | よい連続は、私たちがこの図のことを、4本の線が中央で合流したものではなく、2本の線が重なっているものと知覚しやすいことを示唆するでしょう。

図5.29 | 閉合は、私たちが一連の断片ではなく、完全な円と長方形を知覚することを示唆しています。
学習へのリンク
ゲシュタルト原理の現実世界での例を示したこのビデオ(http://openstax.org/l/gestalt)を見て、さらに学んでください。
ゲシュタルト理論家によれば、パターン知覚、つまり私たちが異なる図や形を区別する能力は、上述の原理に従うことによって生じます。あなたはおそらく、自分の知覚が現実の世界と正確に一致しているとほぼ確信しているでしょうが、必ずしもそうではありません。私たちの知覚は、知覚仮説に基づいています。知覚仮説とは、私たちが感覚情報を解釈する際に行う、知識に裏付けられた推測のことです。それらの仮説は、私たちの人格、経験、および期待を含む、さまざまな要素によって情報が与えられます。私たちはこれらの仮説を使って、知覚的構えを作ります。たとえば、研究では、言葉のプライミングを受けた人が、複雑で曖昧な図形について偏った解釈を生み出すことが示されています(Goolkasian & Woodbury, 2010)。
深く掘り下げてみよう
知覚の深み:偏見、先入観、および文化的要因
この章では、あなたは知覚が複雑なプロセスであることを学んできました。知覚は感覚から組み立てられますが、私たち自身の経験、偏見、先入観、および文化の影響を受け、人によって大きく異なることがあります。研究では、暗黙の人種的な先入観や固定観念が知覚に影響を与えることが示唆されています。たとえば、いくつかの研究では、非黒人の参加者は、武器の画像と黒人の画像が対になっていると、武器をより早く識別し、武器ではないものを武器と識別しがちになることが実証されています(Payne, 2001; Payne, Shimizu, & Jacoby, 2005)。さらに、白人の個人がビデオゲームにおいて武装した標的を撃つかどうかの判断は、標的が黒人の場合には、より素早く行われます(Correll, Park, Judd, & Wittenbrink, 2002; Correll, Urland, & Ito, 2006)。この研究は、過去数十年の間に、若い黒人が殺害され、その殺害した人々が非武装の個人を武装しているものと考えた、および/または、自分の安全に対して何らかの脅威を表していると考えたと主張したという非常に注目度の高い事件が多数発生していることを考慮すると、重要なものです。
重要用語
絶対閾値:その刺激が50%の確率で検出されるために存在しなければならない、刺激エネルギーの最小量
残像:刺激を取り除いた後も視覚的な感覚が継続すること
振幅:波の高さ
基底膜:蝸牛の中にある薄い一片の組織で、聴覚系の感覚受容体として機能する有毛細胞を含む
両耳手がかり:音を定位するための両耳の手がかり
両眼手がかり:両方の目を使用することに依拠する手がかり
両眼視差:それぞれの目がわずかに異なる世界の見え方を受け取ること
盲点:視野の一部で、私たちが視覚情報に反応することができない場所
ボトムアップ処理:感覚入力から知覚を構築するシステム
閉合:知覚を一連の部品としてではなく、完全な物体として整理すること
蝸牛:液体で満たされたカタツムリのような形をした構造物で、聴覚系の感覚受容体細胞を収容する
人工内耳:マイク、音声処理装置、および電極アレイで構成される電子機器で、聴神経を直接刺激して脳に情報を伝達する
伝音性難聴:鼓膜の振動および/または耳小骨の動きがうまくいかないこと
錐体:明るい光の条件下で最もよく機能し、色を感知する特殊な光受容体
先天性難聴:生まれつき耳が聞こえないこと
先天性無痛覚症:痛みを経験することができない遺伝性障害
角膜:眼球を覆う透明な膜
聴覚障害:部分的または完全に聞くことのできない状態
デシベル(dB):音の強さを表す対数の単位
奥行き知覚:奥行きを知覚する能力
電磁スペクトル:私たちの環境で生じるすべての電磁放射
図と地の関係性:私たちの視覚世界を図と地に分けること
中心窩:網膜の小さなくぼみで、錐体を含む
周波数:所与の期間内にある地点を通過する波の数
ゲシュタルト心理学:「全体は部分の総和とは異なる」という考え方に基づいた心理学の分野
よい連続(または、連続):私たちは、ギザギザした折れ線よりも、連続した滑らかな流れのある線を知覚しやすい
有毛細胞:内耳の聴覚受容体細胞
ヘルツ(Hz):1秒あたりのサイクル数で、周波数の尺度
非注意性盲目:注意を払っていないために、完全に見えている何かに気づくことができないこと
砧骨:中耳の耳小骨
炎症性疼痛:あるタイプの組織損傷が発生したことを示す信号
両耳間レベル差:身体の片側からやって来る音が、頭部を通過する際に音波が減衰するため、最も近い耳でより強く聞こえること
両耳間タイミング差:ある音波がそれぞれの耳に到達する時間のわずかな違い
虹彩:眼球の色のついた部分
丁度可知差異:2つの刺激の間の違いを検出するために、必要な刺激の差異
運動感覚:空間における身体の動きの知覚
水晶体:目に入る光の焦点をさらに合わせるための湾曲した透明な構造体
線遠近法:2本の平行線が収束しているように見えるときに、画像に奥行きを知覚すること
槌骨:中耳の耳小骨
マイスナー小体:圧力と低周波数の振動に反応する触覚受容体
メニエール病:内耳の構造が変性し、聴覚の喪失、耳鳴り、めまい、および内耳内の圧力の上昇を引き起こす病気
メルケル盤:軽い接触に反応する触覚受容体
単耳手がかり:音を定位するための片耳の手がかり
単眼手がかり:片方の目のみを必要とする手がかり
神経障害性疼痛:末梢神経系または中枢神経系のニューロンの損傷による痛み
侵害受容:潜在的な危害およびおそらく痛みを示す感覚信号
嗅球:前頭葉の先端にある球状の構造で、嗅神経が始まる場所
嗅覚受容体:嗅覚系の感覚細胞
色覚の反対-処理理論:色は黒色-白色、黄色-青色、赤色-緑色という反対のペアでコード化される
視交叉:脳の腹側面のすぐ下にあるX字型の構造物で、両眼からの視神経が合流し、視野の両側からの情報が脳の反対側に分離される
視神経:網膜からの視覚情報を脳に伝える
パチニ小体:一時的な圧力や高周波数の振動を感知する触覚受容体
パターン知覚:さまざまな図や形を区別する能力
山(または、頂点):波の最高点
知覚:感覚情報が解釈され、意識的に経験される方法
知覚仮説:感覚情報を解釈するために用いられる、知識に裏付けられた推測
フェロモン:他の個体から送られてくる化学的メッセージ
光受容体:光を感知する細胞
耳介:頭部から突き出た目に見える耳の部分
高さ:音の周波数についての知覚
音の高さの知覚についての場所理論:基底膜の異なる部分が異なる周波数の音に対して感受性がある
閉合の原理:知覚を一連の部品としてではなく、完全な物体として整理すること
固有受容感覚:体の位置の知覚
近接:お互いに近くにあるものは一緒にグループ化される傾向がある
瞳孔:光が通過する目の小さな開口部
網膜:光を感知する目の内膜
桿体:光の少ない条件でよく機能する特殊な光受容体
ルフィニ小体:伸縮を感知する触覚受容体
感覚:感覚情報が感覚受容体によって検出されたときに起こるもの
感音性難聴:蝸牛から脳への神経信号の伝達がうまくいかないこと
感覚適応:長期間にわたる比較的一定の刺激を知覚できなくなること
信号検出理論:現在の精神状態に応じた刺激の検出の変化
類同:似ているものは一緒にグループ化される傾向がある
あぶみ骨:中耳の耳小骨
サブリミナル・メッセージ:意識的に認識できる閾値を下回る形で与えられるメッセージ
味蕾:味蕾の中心にある孔に突き出た毛状の味覚受容細胞の集合体
音の高さの知覚についての時間理論:音の周波数は感覚ニューロンの活動レベルによってコード化される
熱受容:温度の知覚
音色:音の純粋さ
トップダウン処理:感覚の解釈は、利用可能な知識、経験、および思考に影響される
伝達:感覚刺激のエネルギーから活動電位への変換
色覚の三色理論:色覚は、3つの錐体グループの活動によって媒介される
谷:波の最低点
鼓膜:鼓膜
うま味:グルタミン酸ナトリウムの味
めまい:回転しているような感覚
前庭感覚:バランスと体の姿勢を維持する能力に貢献する
可視スペクトル:私たちが目で見ることのできる電磁スペクトルの部分
波長:ある頂点から次の頂点までの波の長さ
この章のまとめ
5.1 感覚と知覚
感覚は、感覚受容体が感覚刺激を検出するときに起こります。知覚とは、それらの感覚を整理し、解釈し、意識的に経験することです。すべての感覚系には、絶対閾値と弁別閾値があります。絶対閾値は、50%の確率で検出されるために必要な刺激エネルギーの最小量のことを指し、弁別閾値は、50%の確率で検出されるために必要な刺激エネルギーの差の最小量のことを指します。感覚適応、選択的注意、および信号検出理論は、何が知覚され、何が知覚されないかを説明するのに役立ちます。また、私たちの知覚は、信念、価値観、先入観、文化、および人生経験を含む、さまざまな要因に影響されます。
5.2 波と波長
光も音も、振幅、波長、音色などの物理的な特性を持つ波の形で表現することができます。波長と周波数は反比例の関係にあり、長い波は低い周波数、短い波は高い周波数となります。視覚系では、光波の波長は一般的に色に関連付けられており、振幅は明るさに関連付けられています。聴覚系では、音の周波数は音の高さに関連付けられており、振幅は音の大きさに関連付けられています。
5.3 視覚
光波は角膜を通過して瞳孔から眼球に入ります。目の水晶体は、この光を集束し、像が網膜の中心窩として知られる領域に焦点が合うようにします。中心窩には、高い視力を有し、明るい光の条件下で最もよく機能する錐体があります。桿体は網膜全体に配置されており、薄暗い環境下で最もよく機能します。視覚情報は、視神経を介して目から出ます。それぞれの視野からの情報は、視交叉で反対側の脳に送られます。その後、視覚情報は脳のいくつかの部位を経て、後頭葉に到達し、そこで処理されます。
色の知覚を説明する2つの理論があります。三色理論は、3つの異なる錐体のグループがわずかに異なる波長の光に合わさり、これらの種類の錐体の活動の組み合わせにより、私たちはすべての色を知覚すると主張します。色覚の反対-処理理論は、色は反対のペアで処理されると主張しており、陰性の残像という興味深い現象のことを説明します。私たちは、単眼と両眼の奥行きの手がかりを組み合わせて、奥行きを知覚します。
5.4 聴覚
音波は耳道に入り、鼓膜を振動させます。その振動が耳小骨を動かします。耳小骨が動くと、あぶみ骨が蝸牛の卵円窓を押し、蝸牛内の液体が動きます。その結果、基底膜に埋め込まれた有毛細胞が肥大化し、聴神経を介して脳に神経信号が送られます。
聴覚の重要な要素として、音の高さの知覚と音の定位があります。私たちが音の高さを知覚する能力は、基底膜にある有毛細胞の発火速度と、それらの膜内の位置との両方に基づいています。音の定位については、単耳手がかりと両耳手がかりの両方を使って、環境中のどこから音が発生したかを特定します。
生まれつき耳が聞こえない人もいれば、年齢や遺伝的素因、および/または環境的原因によって耳が聞こえなくなる人もいます。鼓膜の振動または耳小骨の動きがうまくいかないことが原因で起こる難聴を伝音性難聴といいます。また、聴神経のインパルスの脳への伝達がうまくいかない難聴を感音性難聴といいます。
5.5 その他の感覚
味(味覚)と匂い(嗅覚)は、舌の上や鼻の中にある受容体が味や匂いの分子と直接結合し、脳に情報を伝達して処理する化学感覚です。私たちが接触、温度、痛みを知覚する能力は、皮膚や体のさまざまな組織に分布する多数の受容体と自由神経終末によって媒介されます。前庭感覚は、卵形嚢、球形嚢、および半規管の有毛細胞が頭の位置や重力の変化に反応することで、バランス感覚の維持に役立ちます。私たちの固有受容感覚系と運動感覚系は、体の筋肉、関節、腱、および皮膚の伸縮や緊張を検出する受容体を通じて、体の位置や動きについての情報を提供します。
5.6 知覚についてのゲシュタルト原理
ゲシュタルト理論家は、感覚と知覚の分野で非常に大きな影響力を持っています。図と地の関係性、近接または類同によるグループ化、よい連続の法則、閉合などのゲシュタルト原理は、私たちがどのようにして感覚情報を整理しているかを説明するのに使われています。私たちの知覚は無謬のものではなく、それらは偏見、先入観、およびその他の要因によって影響を受けることがあります。
レビュー問題
1.________とは、50%の確率で検出されるために必要な、刺激エネルギーの最小量を指します。
a.絶対閾値
b.弁別閾値
c.丁度可知差異
d.伝達
2.変化しない刺激に対する感度の低下は、________として知られています。
a.伝達
b.弁別閾値
c.感覚適応
d.非注意性盲目
3.________は、感覚刺激のエネルギーを神経インパルスに変換することを含みます。
a.感覚適応
b.非注意性盲目
c.弁別閾値
d.伝達
4.________は、感覚情報が整理され、解釈され、意識的に経験されるときに起こります。
a.感覚
b.知覚
c.伝達
d.感覚適応
5.短波長から長波長へと移行する際の色の知覚のパターンと正しく一致するのは、以下のうちどれですか?
a.赤色→橙色→黄色
b.黄色→橙色→赤色
c.黄色→赤色→橙色
d.橙色→黄色→赤色
6.可視スペクトルには、約________の範囲の光が含まれています。
a.400~700nm
b.200~900nm
c.20~20000Hz
d.10~20dB
7.電磁スペクトルには、________が含まれます。
a.電波
b.X線
c.赤外光
d.上記のすべて
8.人間にとっての可聴域は、________です。
a.380~740Hz
b.10~20dB
c.300dB未満
d.20~20000Hz
9.音波の周波数、振幅、タイミングによって影響される音の質は、________として知られています。
a.高さ
b.音調
c.電磁
d.音色
10.________は、網膜の小さなくぼみで、錐体を含んでいます。
a.視交叉
b.視神経
c.中心窩
d.虹彩
11.________は、明るい光の条件下で最もよく機能します。
a.錐体
b.桿体
c.網膜神経節細胞
d.線条皮質
12.________の奥行きの手がかりは、両目を使う必要があります。
a.単眼
b.両眼
c.線遠近法
d.協調
13.もしあなたが緑の丸を比較的長い時間見つめた後、真っ白なスクリーンに視線を移すと、あなたは________の陰性の残像が見えるでしょう。
a.青色
b.黄色
c.黒色
d.赤色
14.基底膜の基部付近に位置する有毛細胞は、________の音に最もよく反応します。
a.低い周波数
b.高い周波数
c.低い振幅
d.高い振幅
15.中耳の3つの耳小骨は、________として知られています。
a.槌骨、砧骨、およびあぶみ骨
b.槌骨、砧骨、およびあぶみ骨
c.耳介、蝸牛、および卵形嚢
d.aとbの両方
16.補聴器は、________の治療に有効かもしれません。
a.メニエール病
b.感音性難聴
c.伝音性難聴
d.両耳間タイミング差
17.2つの耳を必要とする手がかりは、________手がかりと呼ばれます。
a.単眼
b.単耳
c.両眼
d.両耳
18.生殖状態について何らかのことを伝えるために、種の2つのメンバーの間でしばしば送られる化学的メッセージは、________と呼ばれます。
a.ホルモン
b.フェロモン
c.メルケル盤
d.マイスナー小体
19.グルタミン酸ナトリウムに関連する味はどれですか?
a.甘味
b.苦味
c.うま味
d.酸味
20.________は、温度や痛みの刺激に対する感覚受容体として機能します。
a.自由神経終末
b.パチニ小体
c.ルフィニ小体
d.マイスナー小体
21.バランスと体の姿勢の維持に関わっているのは、以下のうちどれですか?
a.聴神経
b.侵害受容器
c.嗅球
d.前庭系
22.________の原則によると、お互いに近くにある物体は一緒にグループ化される傾向があります。
a.類同
b.よい連続
c.近接
d.閉合
23.物事を一連の部品としてではなく、完全な物体として知覚する私たちの傾向は、________の原則として知られています。
a.閉合
b.よい連続
c.近接
d.類同
24.________の法則によると、私たちは途切れ途切れの線やギザギザの線よりも、滑らかに流れる線を知覚する傾向があります。
a.閉合
b.よい連続
c.近接
d.類同
25.視覚的な表示における主な焦点は、________として知られています。
a.閉合
b.知覚的構え
c.地
d.図
批判的思考の問題
26.感覚されたものがすべて知覚されるわけではありません。あなたは、何かが感覚されることなく知覚される場合があると思いますか?
27.刺激の強さに応じて丁度可知差異がどのように変化するかということについての新規な例を出してください。
28.他の生物種が人間と比較して、視覚と聴覚の両方の刺激に対して非常に異なる感度の範囲を有するのはなぜだと考えますか?
29.人間が可聴域の中間部分に位置する周波数の音に特に敏感なのはなぜだと考えますか?
30.色の知覚に関する2つの理論を比較してみてください。両者は全く違うものなのでしょうか?
31.色は私たちの環境の物理的特性ではありません。あなたは、色覚には(もしあるとしたら)どのような機能があると思いますか?
32.あなたが音の定位について読んだことを踏まえて、進化の観点から、音の定位は生存をどのように促進するでしょうか?
33.周波数4000Hzまでの音波の高さを知覚する能力を説明するのに、どうして時間理論と場所理論の両方を使うことができるのでしょうか?
34.多くの人が車、飛行機、または船で移動中に吐き気を催します。あなたはこれを感覚的な相互作用の機能としてどのように説明できるでしょうか?
35.もし、誰かが「大きな怪我に伴う痛みを感じないためならば何でもする」と言っているのを聞いたとしたら、ここまで読んできた内容を踏まえて、あなたはどう答えますか?
36.あなたは、女性の痛みの感じ方は、男性とは違うと思いますか?それはなぜだと考えますか?
37.ゲシュタルト心理学の中心的な教義は、「全体は部分の総和とは異なる」というものです。これは、知覚の文脈においてどのような意味を持ちますか?
38.以下の図を見てください。人々がアヒルを見るかウサギを見るかということに対して、あなたはどのような影響を与えることができるでしょうか?
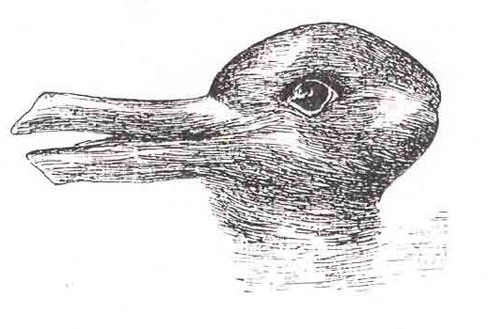
図5.30
個人的に当てはめてみる問題
39.あなたの注意が他に集中していたために、周りの何かに気づかなかったときのことを考えてみてください。誰かがそれを指摘したとき、あなたはすぐに気づかなかったことに驚きましたか?
40.もしあなたが家族の一員としてペットを飼っていたならば、ペットはあなたが聞こえないような音をしばしば聞いているようだということに気付いたでしょう。あなたはこの章を読み終わりましたが、その理由についておそらく何らかの洞察を有しています。このような授業を受ける機会がなかった友人に対して、そのことをどのように説明しますか?
41.あなたの写真や個人的な芸術作品をいくつか見てみてください。線遠近法が奥行きの手がかりになるような例を見つけられますか?
42.もしあなたが、視力と聴力のどちらかを失うことを選択しなければいけないとしたら、どちらを選びますか?その理由は何ですか?
43.先に述べたように、食べ物の風味は味覚と嗅覚の両方の情報の相互作用を表しています。あなたが風邪やインフルエンザのために鼻づまりがひどかったときのことを考えてみてください。その時に食べた食べ物の風味に何らかの変化があったことに気づきましたか?
44.あなたがラジオである曲を聴いて一緒に歌っていたのに、後で間違った歌詞を歌っていたことに気付いたことはありますか?正しい歌詞を見つけた後、その曲に対するあなたの知覚は変わりましたか?
参考文献
Aaron, J. I., Mela, D. J., & Evans, R. E. (1994). The influences of attitudes, beliefs, and label information on perceptions of reduced-fat spread. Appetite, 22, 25–37.
Abraira, V. E., & Ginty, D. D. (2013). The sensory neurons of touch. Neuron, 79, 618–639.
Animals in science/alternatives. NEAVS. https://www.navs.org/what-we-do/keep-you-informed/science-corner/alternatives/alternatives-to-animal-testing/#.XmsJ1qhKhPY
Ayabe-Kanamura, S., Saito, S., Distel, H., Martínez-Gómez, M., & Hudson, R. (1998). Differences and similarities in the perception of everyday odors: A Japanese-German cross-cultural study. Annals of the New York Academy of Sciences, 855, 694–700.
Birch, J. (2012). Worldwide prevalence of red-green color deficiency. Journal of the Optical Society of America A, 29, 313–320. https://www.osapublishing.org/josaa/abstract.cfm?uri=josaa-29-3-313
Chen, Q., Deng, H., Brauth, S. E., Ding, L., & Tang, Y. (2012). Reduced performance of prey targeting in pit vipers with contralaterally occluded infrared and visual senses. PloS ONE, 7(5), e34989. doi:10.1371/journal.pone.0034989
Comfort, A. (1971). Likelihood of human pheromones. Nature, 230, 432–479.
Correll, J., Park, B., Judd, C. M., & Wittenbrink, B. (2002). The police officer’s dilemma: Using ethnicity to disambiguate potentially threatening individuals. Journal of Personality and Social Psychology, 83, 1314–1329.
Correll, J., Urland, G. R., & Ito, T. A. (2006). Event-related potentials and the decision to shoot: The role of threat perception and cognitive control. The Journal of Experimental Social Psychology, 42, 120–128.
Dunkle T. (1982). The sound of silence. Science, 82, 30–33.
Egeth, H., & Yantis, S. (1997). Visual attention: Control, representation, and time course. Annual Review of Psychology, 48, 269–297. https://www.researchgate.net/publication/14163796_Visual_Attention_Control_Representation_and_Time_Course
Fawcett, S. L., Wang, Y., & Birch, E. E. (2005). The critical period for susceptibility of human stereopsis. Investigative Ophthalmology and Visual Science, 46, 521–525.
Fine, M. S., & Minnery, B. S. (2009). Visual salience affects performance in a working memory task. Journal of Neuroscience, 29, 8016–8021. https://www.jneurosci.org/content/29/25/8016
Furlow, F. B. (1996, 2012). The smell of love. http://www.psychologytoday.com/articles/200910/the-smell-love
Galanter, E. (1962). Contemporary Psychophysics. In R. Brown, E.Galanter, E. H. Hess, & G. Mandler (Eds.), New directions in psychology. Holt, Rinehart & Winston.
Garland, E. L. (2012). Pain processing in the human nervous system: A selective review of nociceptive and biobehavioral pathways. Primary Care, 39, 561–571.
Goolkasian, P. & Woodbury, C. (2010). Priming effects with ambiguous figures. Attention, Perception & Psychophysics, 72, 168–178.
Grothe, B., Pecka, M., & McAlpine, D. (2010). Mechanisms of sound localization in mammals. Physiological Reviews, 90, 983–1012.
Hartline, P. H., Kass, L., & Loop, M. S. (1978). Merging of modalities in the optic tectum: Infrared and visual integration in rattlesnakes. Science, 199, 1225–1229.
Hubel, D. H., & Wiesel, T. N. (1959). Receptive fields of single neurones in the cat’s striate cortex. Journal of Physiology, 148, 574–591. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1363130/
Hubel, D. H., & Wiesel, T. N. (1962). Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. Journal of Physiology, 160, 106–154. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1359523/
Hubel, D. H., & Weisel, T. N. (1963). Single-cell responses in striate cortex of kittens deprived of vision in one eye. Journal of Neurophysiology. https://journals.physiology.org/doi/abs/10.1152/jn.1963.26.6.1003
Hubel, D. H., & Wiesel, T. N. (1970). The period of susceptibility to the physiological effects of unilateral eye closure in kittens. The Journal of Physiology, 206(2). https://physoc.onlinelibrary.wiley.com/doi/abs/10.1113/jphysiol.1970.sp009022
Kaiser, P. K. (1997). The joy of visual perception: A web book. <http://www.yorku.ca/eye/noframes.htmL
Khan, S., & Chang, R. (2013). Anatomy of the vestibular system: A review. NeuroRehabilitation, 32, 437–443.
Kinnamon, S. C., & Vandenbeuch, A. (2009). Receptors and transduction of umami taste stimuli. Annals of the New York Academy of Sciences, 1170, 55–59.
Kujawa, S. G., & Liberman, M. C. (2006). Acceleration of age-related hearing loss by early noise exposure: Evidence of a misspent youth. Journal of Neuroscience, 26, 2115–2123. https://www.jneurosci.org/content/26/7/2115
Kunst-Wilson, W. R., & Zajonc, R. B. (1980). Affective discrimination of stimuli that cannot be recognized. Science, 207, 557–558.
Lackner, J. R., & DiZio, P. (2005). Vestibular, proprioceptive, and haptic contributions to spatial orientation. Annual Review of Psychology, 56, 115–147.
Land, E. H. (1959). Color vision and the natural image. Part 1. Proceedings of the National Academy of Science, 45(1), 115–129.
Latham, S. (2012). U.S. law and animal experimentation: A critical primer. The Hastings Center. http://animalresearch.thehastingscenter.org/report/u-s-law-and-animal-experimentation-a-critical-primer/
Le, T. N., Straatman, L. V., Lea, J., & Westerberg, B. (2017). Current insights in noise-induced hearing loss: A literature review of the underlying mechanism, pathophysiology, asymmetry, and management options. Journal of Otolaryngology - Head & Neck Surgery, 46(1), 41. https://journalotohns.biomedcentral.com/articles/10.1186/s40463-017-0219-x
Liem, D. G., Westerbeek, A., Wolterink, S., Kok, F. J., & de Graaf, C. (2004). Sour taste preferences of children relate to preference for novel and intense stimuli. Chemical Senses, 29, 713–720.
Lodovichi, C., & Belluscio, L. (2012). Odorant receptors in the formation of olfactory bulb circuitry. Physiology, 27, 200–212.
Loersch, C., Durso, G. R. O., & Petty, R. E. (2013). Vicissitudes of desire: A matching mechanism for subliminal persuasion. Social Psychological and Personality Science, 4(5), 624–631.
Mack, A., & Rock, I. (1998). Inattentional Blindness. MIT Press.
Maffei, A., Haley, M., & Fontanini, A. (2012). Neural processing of gustatory information in insular circuits. Current Opinion in Neurobiology, 22, 709–716.
Miller, B. T., & D’Esposito, M. (2005). Searching for “the top” in top-down control. Neuron, 48, 535–538. https://www.sciencedirect.com/science/article/pii/S0896627305009360
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annual Review of Neuroscience, 24, 167–202. https://www.annualreviews.org/doi/abs/10.1146/annurev.neuro.24.1.167
Milner, A. D., & Goodale, M. A. (2008). Two visual systems re-viewed. Neuropsychological, 46, 774–785.
Mizushige, T., Inoue, K., Fushiki, T. (2007). Why is fat so tasty? Chemical reception of fatty acid on the tongue. Journal of Nutritional Science and Vitaminology, 53, 1–4.
Most, S. B., Simons, D. J., Scholl, B. J., & Chabris, C. F. (2000). Sustained inattentional blindness: The role of location in the detection of unexpected dynamic events. PSYCHE, 6(14).
Nelson, M. R. (2008). The hidden persuaders: Then and now. Journal of Advertising, 37(1), 113–126.
Niimura, Y., & Nei, M. (2007). Extensive gains and losses of olfactory receptor genes in mammalian evolution. PLoS ONE, 2, e708.
Okawa, H., & Sampath, A. P. (2007). Optimization of single-photon response transmission at the rod-to-rod bipolar synapse. Physiology, 22, 279–286.
Payne, B. K. (2001). Prejudice and perception: The role of automatic and controlled processes in misperceiving a weapon. Journal of Personality and Social Psychology, 81, 181–192.
Payne, B. K., Shimizu, Y., & Jacoby, L. L. (2005). Mental control and visual illusions: Toward explaining race-biased weapon misidentifications. Journal of Experimental Social Psychology, 41, 36–47.
Peck, M. (2012, July 19). How a movie changed one man’s vision forever. http://www.bbc.com/future/story/20120719-awoken-from-a-2d-world
Peterson, M. A., & Gibson, B. S. (1994). Must figure-ground organization precede object recognition? An assumption in peril. Psychological Science, 5, 253–259.
Petho, G., & Reeh, P. W. (2012). Sensory and signaling mechanisms of bradykinin, eicosanoids, platelet-activating factor, and nitric oxide in peripheral nociceptors. Physiological Reviews, 92, 1699–1775.
Proske, U. (2006). Kinesthesia: The role of muscle receptors. Muscle & Nerve, 34, 545–558.
Proske, U., & Gandevia, S. C. (2012). The proprioceptive senses: Their roles in signaling body shape, body position and movement, and muscle force. Physiological Reviews, 92, 1651–1697.
Purvis, K., & Haynes, N. B. (1972). The effect of female rat proximity on the reproductive system of male rats. Physiology & Behavior, 9, 401–407.
Radel, R., Sarrazin, P., Legrain, P., & Gobancé, L. (2009). Subliminal priming of motivational orientation in educational settings: Effect on academic performance moderated by mindfulness. Journal of Research in Personality, 43(4), 1–18.
Rauschecker, J. P., & Tian, B. (2000). Mechanisms and streams for processing “what” and “where” in auditory cortex. Proceedings of the National Academy of Sciences, USA, 97, 11800–11806.
Renier, L. A., Anurova, I., De Volder, A. G., Carlson, S., VanMeter, J., & Rauschecker, J. P. (2009). Multisensory integration of sounds and vibrotactile stimuli in processing streams for “what” and “where.” Journal of Neuroscience, 29, 10950–10960.
Rensink, R. A. (2004). Visual sensing without seeing. Psychological Science, 15, 27–32.
Rock, I., & Palmer, S. (1990). The legacy of Gestalt psychology. Scientific American, 262, 84–90.
Roper, S. D. (2013). Taste buds as peripheral chemosensory receptors. Seminars in Cell & Developmental Biology, 24, 71–79.
Russell, M. J. (1976). Human olfactory communication. Nature, 260, 520–522.
Sachs, B. D. (1997). Erection evoked in male rats by airborne scent from estrous females. Physiology & Behavior, 62, 921–924.
Segall, M. H., Campbell, D. T., & Herskovits, M. J. (1963). Cultural differences in the perception of geometric illusions. Science, 139, 769–771.
Segall, M. H., Campbell, D. T., & Herskovits, M. J. (1966). The influence of culture on visual perception. Bobbs-Merrill.
Segall, M. H., Dasen, P. P., Berry, J. W., & Poortinga, Y. H. (1999). Human behavior in global perspective (2nd ed.). Allyn & Bacon.
Semaan, M. T., & Megerian, C. A. (2010). Contemporary perspectives on the pathophysiology of Meniere’s disease: implications for treatment. Current opinion in Otolaryngology & Head and Neck Surgery, 18(5), 392–398.
Shamma, S. (2001). On the role of space and time in auditory processing. Trends in Cognitive Sciences, 5, 340–348.
Simons, D. J., & Chabris, C. F. (1999). Gorillas in our midst: Sustained inattentional blindness for dynamic events. Perception, 28, 1059–1074.
Spors, H., Albeanu, D. F., Murthy, V. N., Rinberg, D., Uchida, N., Wachowiak, M., & Friedrich, R. W. (2013). Illuminating vertebrate olfactory processing. Journal of Neuroscience, 32, 14102–14108.
Spray, D. C. (1986). Cutaneous temperature receptors. Annual Review of Physiology, 48, 625–638.
Strain, G. M. (2003). How well do dogs and other animals hear? http://www.lsu.edu/deafness/HearingRange.html
Swets, J. A. (1964). Signal detection and recognition by human observers. Psychological Bulletin, 60, 429–441.
Ungerleider, L. G., & Haxby, J. V. (1994). ‘What’ and ‘where’ in the human brain. Current Opinion in Neurobiology, 4, 157–165.
U.S. Department of Health and Human Services, National Institutes of Health, & National Institute on Deafness and Other Communication Disorders (NIDCD). (2014). Noise-induced hearing loss. NIH Publication No. 14-4233. https://www.nidcd.nih.gov/health/noise-induced-hearing-loss
U.S. National Library of Medicine. (2013). Genetics home reference: Congenital insensitivity to pain. http://ghr.nlm.nih.gov/condition/congenital-insensitivity-to-pain
Vecera, S. P., & O’Reilly, R. C. (1998). Figure-ground organization and object recognition processes: An interactive account. Journal of Experimental Psychology-Human Perception and Performance, 24, 441–462.
Wakakuwa, M., Stavenga, D. G., & Arikawa, K. (2007). Spectral organization of ommatidia in flower-visiting insects. Photochemistry and Photobiology, 83, 27–34.
Weisel, T. N., & Hubel, D. H. (1963). Single-cell responses in striate cortext of kittens deprived of vision in one eye. Journal of Neurophysiology. https://journals.physiology.org/doi/abs/10.1152/jn.1963.26.6.1003
Weller, A. (1998). Human pheromones: Communication through body odour. Nature, 392, 126–127.
Wells, D. L. (2010). Domestic dogs and human health: An overview. British Journal of Health Psychology, 12, 145–156.
Wolfgang-Kimball, D. (1992). Pheromones in humans: myth or reality?. http://www.anapsid.org/pheromones.html
Wysocki, C. J., & Preti, G. (2004). Facts, fallacies, fears, and frustrations with human pheromones. The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology, 281, 1201–1211.
Yantis, S., & Egeth, H. (1999). On the distinction between visual salience and stimulus-driven attentional capture. Journal of Experimental Psychology: Human Perception and Performance, 25, 661–676. https://psycnet.apa.org/doiLanding?doi=10.1037%2F0096-1523.25.3.661
この訳文は元の本のCreative Commons BY 4.0ライセンスに従って同ライセンスにて公開します。問題がありましたら、可能な限り早く対応いたしますので、ご連絡ください。また、誤訳・不適切な表現等ありましたらご指摘ください。この本は、https://openstax.org/details/books/psychology-2eで無料でダウンロードできます。